
Remember me
HPLC analysis of G. mellonella hemolymph provided insights into the presence and quantification of various bioactive compounds with antioxidative and anti-inflammatory properties. These include quinic acid, ellagic acid, cinnamic chlorogenic acid and resorcinol, enanthic acid, butyric acid, pentadecanoic acid, myristoleic acid, heneicosanoic acid, arachidonic acid, stearic acid, oleic acid, linoleic acid, eicosatrienoic acid, and eicosapentaenoic acid.
The hemolymph was found to have a very low level of toxicity and did not cause any mortality exceeding 10,000 mg/kg body weight. On the basis of the acute toxicity studies shown in Fig. 1, the LD50 of the hemolymph is estimated to be greater than 3000 mg/kg b wt (highest concentration without any signs). An acceptable daily intake (ADI) dose of 300 mg/kg b wt was estimated (1/10 LD50) and applied to test its effect on arthritis-induced rats.
Fig. 1
Acute toxicity of hemolymph illustrated the correlation between administrated doses (mg/kg BW) and toxicity signs
PICKLE/HUSCH database predictionFigure 2A displays the CRP/OST/PADI4 interactions that were explored with other genes and with each other. This information was obtained from the PICKLE database. The interaction between CRP, OST, and arthritis, specifically involving PADI4 (Peptidyl Arginine Deiminase 4), is an area of research focused on understanding the pathogenesis of arthritis. Hence, CRP is an acute-phase protein associated with inflammation, OST, a bone-derived protein that particularly plays a key role in bone remodeling processes related to arthritis, and PADI4, an enzyme involved in the posttranslational modification of proteins by citrullination, has been linked to autoimmune responses in arthritis, as citrullinated proteins are often targeted by the immune system in rheumatoid arthritis. The current in-silico simulation of their interaction revealed complex and ongoing crosstalk mechanisms and pathways that modulate the development and progression of arthritis. By predicting the interactions of CRP and OST with PADI4, a key arthritis-associated gene, PICKLE helps elucidate how these proteins contribute to inflammatory pathways. This finding directly correlates with the in-vivo evaluation of osteocalcin’s role in joint inflammation and bone remodeling.
Fig. 2
A Concentric layout network visualization of CRP/OST/PADI4 interaction. Data retrieved from PICKLE (Accessed on 3 March 2024). The concentric layout displays 54 nodes (proteins) and 139 edges (interactions), with red nodes highlighting the central proteins of interest (CRP, OST, and PADI4) and green nodes representing first-neighbor interacting proteins. The edges depict known protein–protein interactions, derived from high-quality experimental and computational sources (PPI quality level 2). Self-loops indicate proteins with autointeractions. The network was normalized at the protein level (UniProt) and filtered using the default cross-checking method. The network structure reveals highly interconnected hub proteins, which may indicate potential regulatory roles in signaling pathways. Highly interconnected nodes, suggests functional relationships and co-regulation of biological pathways; Self-looping proteins, indicate proteins with autointeractions or self-regulatory functions; Cross-talk between pathways, CRP, OST, and PADI4 may bridge different biological processes, influencing inflammation, immune response, and protein modifications. This network can help identify potential therapeutic targets and guide further studies into inflammatory disease mechanisms. B Bone tissue specific cells annotation, data retrieved from HUSCH, comparative gene exploration for BGLAP, IL-6, TNF-α, and PADI4 expression in bone tissue. C Adipose stromal specific cells annotation, data retrieved from HUSCH, comparative gene exploration for BGLAP, IL-6, TNF-α, and PADI4 expression in adipose stromal tissue
Figures 2B & C present cell annotations and an examination of the expression of BGLAP (osteocalcin), IL-6, TNF-α, and PADI4 in arthritis-relevant tissues, which provide insights into their involvement in RA pathophysiology. This finding aligns with the in-vivo assessment of cytokine levels and miR-146a regulation, reinforcing the molecular link between systemic inflammation and bone metabolism.
Annotation of cell types in bone tissue (dataset: HU_0370_Bone_GSE169396) is shown in Fig. 2B. The breakdown of observational BGLAP (osteocalcin), IL-6, PADI4, and TNF-α expression levels revealed high expression of IL-6, TNF-α, and PADI4 in monocytes/macrophages, supporting their central role in driving inflammation and immune activation in arthritis. These cytokines contribute to osteoclast activation, leading to bone resorption. The exclusive expression of BGLAP in osteoblasts/osteocytes highlights its involvement in bone matrix production and integrity, which may be disrupted by inflammatory cytokines such as IL-6 and TNF-α.
Annotation of cell types in adipose stromal tissues is shown in Fig. 2C. The chart highlights the expression patterns of BGLAP, IL-6, PADI4, and TNF-α. Elevated expression of IL-6 and TNF-α in pericytes and smooth muscle cells indicates their involvement in inflammatory signaling and tissue remodeling within adipose stromal tissues, which may contribute to arthritis-associated inflammation. Minimal or negligible expressions of BGLAP suggested a limited role in adipose stromal tissues compared with bone-related contexts.
Gene–gene interactions and pathwaysThe CRP, OST (BGLAP), and PADI4 gene‒gene interactions and pathways, as compiled from curated databases and text mining at the UCSC Genomics Institute, are shown in Fig. 3. The interactions and pathways involving these genes are of interest for understanding various biological processes, particularly in the context of arthritis as an inflammatory disease. Figure 3 provides a brief overview of these interactions and pathways, which are likely complex and multifaceted, contributing to various aspects of inflammation, bone metabolism, and autoimmune responses involved in arthritis. Identifying the top interacting genes for CRP, BGLAP, and PADI4 allows for a deeper understanding of their regulatory networks. This bioinformatics analysis supports the in-vivo exploration of how miR-146a modulates inflammatory signaling pathways, particularly the IL-6 and TNF-α pathways, in RA progression. Studying these interactions and pathways can provide insights into disease mechanisms and potential therapeutic targets for managing inflammatory and autoimmune conditions.
Fig. 3
CRP/OST(BGLAP)/PADI4 top interacting genes. Data retrieved from UCSC Genome Browser Gene Interaction Graph, highlighting the Drug Bank interaction, obtained via gene interactions and pathways. Accessed on 3 March 2024. CRP: chr1:159682078–159684379; BGLAP: chr1:156211950–156213123; PADI4: chr1:17634689–17690495. Interactions are colored by support: Black colored genes: treatment hits by Drug Bank; Grey: interaction from several datasets with only text mining; light blue: interaction database; blue: pathway database; Dashed lines indicate interactions without text mining support
These in-silico predictions are corroborated by in-vivo results, offering valuable insights into the molecular mechanisms underlying RA pathogenesis and the potential therapeutic effects of G. mellonella hemolymph. This integrative approach, encompassing computational molecular modeling and in-vivo validation, underscores the significance of these interactions in the development of novel antirheumatic treatments.
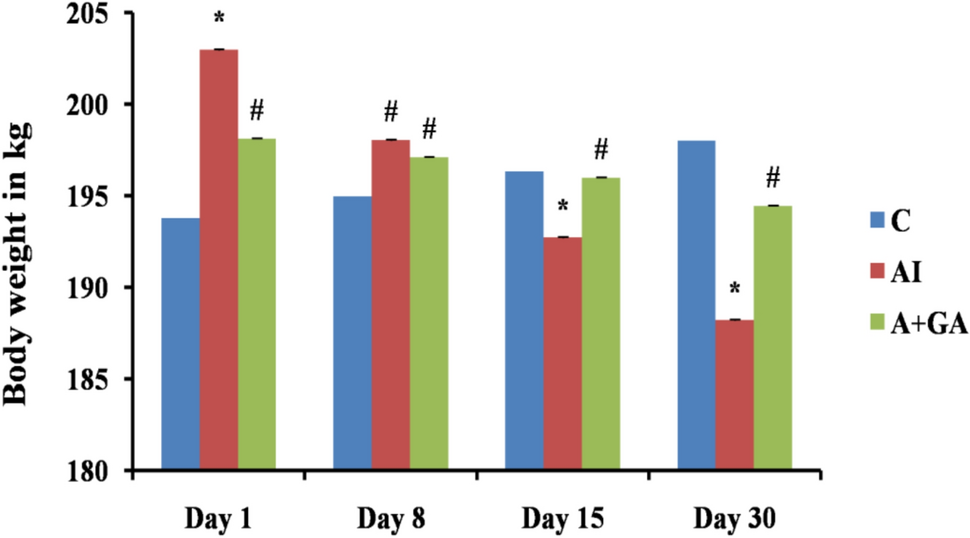
Body and relative organ weightsCompared with control rats, rheumatoid arthritis and RA + MTX rats presented highly significant reductions (p < 0.01) in terms of body weight gain (Table 1). Compared with control rats, hemolymph arthritis-treated rats presented a slight decrease in body weight gain that was not statistically significant but was significantly greater (p < 0.01) than that of the RA + MTX group.
Table 1 The body and relative organ weightThe liver, kidney, and heart relative weights of the RA and RA + MTX rats were not significantly greater than those of the control group. The same result was observed for the relative weight of the spleen in the RA + MTX group. The relative spleen weight in the arthritic group was not significantly lower (p < 0.05) than that in the control group (Table 1).
Similarly, when hemolymph extract was administered, the relative liver weight dramatically increased (p < 0.05). Additionally, a nonsignificant increase in the heart and spleen relative weights and a nonsignificant reduction in the kidney relative weight were observed relative to those in the control group (Table 1).
Inflammatory infiltratesFigure 4A shows the paw diameter change (cm) in the different studied groups as an indicator of inflammatory infiltration, which is typically perivascular expression. Edema in the arthritic paw refers to the accumulation of inflammatory cells within the paw, accompanied by edema or swelling. Compared with the control group (0.0840 ± 0.01), the rheumatoid arthritis group (RA) presented a highly significant increase (p < 0.01) in the mean paw diameter (0.272 ± 0.08), whereas both the methotrexate (RA + MTX)- and hemolymph-treated groups (RA + GHE) presented nonsignificant changes (0.120 ± 0.04), whereas the G. mellonella hemolymph-Arthritis group (RA + GHE) presented a nonsignificant decrease (0.066 ± 0.01) in paw diameter. These results suggest that G. mellonella hemolymph may have a mitigating effect on arthritis comparable to that of MTX.
Fig. 4
A Change in the paw diameter (cm) represented as (mean ± SE) in different experimental groups. Control group (C), Rheumatoid arthritis group (RA), Methotrexate arthritic treated group (RA + M), and hemolymph arthritic treated group (RA + GHE). The mean difference is significant at p < 0.05. a: significant vs control, b: significant vs RA group. B Drawn pathway maps of synovial angiogenesis and leukocyte infiltration, obtained via KEGG PATHWAY Database, accessed 25 November 2024
Abnormal activation of the immune system elevates proinflammatory cytokine levels, which can promote synovial angiogenesis and leukocyte infiltration. The synovium forms a hyperplastic pannus with infiltrated macrophage-like and fibroblast-like synoviocytes and invades joints by secreting proteinases and inducing osteoclast differentiation. These data underscore the cell-specific expression of inflammatory mediators and bone-related markers, offering insights into tissue-specific regulatory mechanisms in arthritis (Fig. 4B).
Hematological and biochemical findingsTable 2 shows the hematological and biochemical changes. Compared with those in the control group, the average Hb and PLT values were significantly lower (p < 0.001) and RBC counts (p < 0.005) in the RA group. However, the WBC value highly significantly increased (P < 0.001).
Table 2 Hematological and biochemical modulatorsCompared with the control group, the RA + MTX group presented markedly lower (p < 0.001) PLTs and WBCs. Additionally, the Hb and RBC values were significantly lower (P < 0.05). In contrast, compared with the RA group, the RA + M group presented significant increases (p < 0.05) in Hb and RBCs.
The hemolymph treatment showed remarkable improvements when compared to the rheumatoid arthritis group. There was a substantial decrease (p < 0.001) in WBCs and a considerable increase (p < 0.001) in Hb and PLTs. Additionally, there was a significant increase (p < 0.05) in RBCs. Furthermore, the PLTs and WBCs were significantly greater (p < 0.0001) in RA + GHE-treated rats than in those treated with MTX.
Regarding the liver enzymes, Table 2 shows a highly significant increase (p < 0.001) in the serum ALT level in both the RA and RA + GHE groups compared with the control group. Moreover, the ALT level was significantly lower (p < 0.001) in the RA + GHE group than in the RA + MTX group.
Fortunately, serum AST levels exhibited a very significant increase (p < 0.001) among all studied groups, as compared to the control group. Moreover, RA + M and RA + GHE resulted in very significant reductions (p < 0.001) compared with those of the rheumatoid arthritis group, highlighting the preservation of the RA + GHE group, which presented the greatest reduction in values.
Similarly, the serum ALP levels were significantly elevated (p < 0.001) in all the experimental groups compared with those in the control group. Compared with the arthritis group, the MTX and hemolymph groups presented significantly lower ALP levels (p < 0.01 and p < 0.001, respectively). Additionally, compared with RA + MTX, G. mellonella hemolymph significantly decreased ALP levels (p < 0.05).
Compared with those in the control group, the serum creatinine and urea levels were significantly elevated (p < 0.001) in all the studied groups, whereas the MTX- and GHE-treated groups presented highly significant reductions (p < 0.001) compared with the RA groups. Compared with MTX-treated rats, GHE-treated rats presented highly significant reductions (p < 0.001) in creatinine and urea levels.
Oxidative biomarkersCompared with those in the control group, the serum levels of TAC in the different experimental groups were significantly lower (p < 0.001) than that of the control group (Table 3). However, compared with those in RA and RA + MTX groups, the mean values of TAC in the hemolymph in the experimental groups were significantly greater (p < 0.001).
Table 3 Oxidative and inflammatory mediators in different experimental groups. Data expressed as Mean ± SDCompared with those in the control group, hepatic MDA and SOD were significantly increased (p < 0.001) in all the experimental groups. However, hemolymph administration significantly reduced the hepatic levels of both MDA and SOD (p < 0.001) compared with those in the untreated and methotrexate-treated arthritis groups.
Inflammatory mediatorsThe data in Table 3 revealed significant elevations (p < 0.001) in the serum levels of CRP and ACCP in the three arthritic groups, either untreated or treated, compared with those in the control group. Compared with RA and RA + MTX, G. mellonella hemolymph administration to arthritic rats significantly (p < 0.001) reduced the serum levels of CRP and ACCP.
The serum levels of IL-6 and TNF‐α, compared with those in the control group, were markedly significantly increased (p < 0.001) in all the studied groups. Moreover, the RA + MTX group presented highly significant reductions (p < 0.001) in the levels of IL-6 and TNF-α compared with those in the RA group. Additionally, the levels of both inflammatory mediators were markedly lower (p < 0.001) in the G. mellonella hemolymph-treated group than in the untreated and RA + MTX groups.
The data in Table 3 show a highly significant increase (p < 0.001) in the miR-146a expression level in the arthritic (RA) group compared with the normal control group. Although both the MTX- and G. mellonella hemolymph-treated groups presented highly statistically significant reductions (p < 0.001) in the miR-146a level, compared with those in the untreated arthritic group, the miR-146a level in the MTX-treated group was still significantly greater (p < 0.001 & p < 0.004) than that in the control group. Moreover, RA + GHE presented markedly lower (p < 0.001) miR-146a expression levels than RA + MTX-treated rats did.
As shown in Figs. 5A & B, a significant positive correlation was observed between miR-146a and both IL-6 (r = 982, p < 0.001) and TNF-α (r = 978, p < 0.001).
Fig. 5
Correlation of miR-146a with IL-6 and TNF-α. A statistically significant positive correlation between miR-146a expression level and IL-6 (r = 0.982, p < 0.001). B IL-6 genes relative adipose tissue expression heatmap, a part of cluster 7 with confidence 1. C IL-6 genes relative connective tissue expression, a part of cluster 62 with confidence 1. D statistically significant positive correlation between miR-146a expression level and TNF-α (r = 0.978, p < 0.001). E TNF-α genes relative adipose tissue expression heatmap, a part of cluster 81 with confidence 1. F TNF-α genes relative connective tissue expression heatmap, a part of cluster 50 with confidence 1. N.B. relative expression from 0.0 to 1.0 (white to dark red color)
Histopathological and immunohistochemicalHistological analysis of H&E-stained sections from the metatarsal joint region in the control group revealed a thin cartilaginous cap composed of actively dividing chondrocytes, either singly or in pairs within lacunae (Fig. 6). This cap overlay a structurally normal cortical bone, characterized by bone lamellae and osteocytes within lacunae. Additionally, a smooth, continuous endosteal surface covered by osteoprogenitor and osteoblast cells was observed. The synovial membrane, fibrohyaline joint capsule, and surrounding hypodermal tissue appeared normal, without any inflammatory or degenerative changes. No exudative or transudative fluids were detected in bone marrow regions rich in hemopoietic cells (Fig. 6A).
Figs. 6
Histological manifestations from meta-tarsal joints of different experimental groups. A normal control group showing cortical bone having bone lamellae and osteocytes inside their lacunae (yellow arrows). The synovial membrane (black arrows), fibro hyaline joint capsule and the surrounding hypodermal tissue appear normal and free from any inflammatory or degenerative changes (blue arrows). B Arthritis group showing damage to all the joint compartments, surrounding tissues and represented by subchondral bone damage, and changes in the synovium that manifest as episodic synovitis. The bone tissue shows osteolytic degeneration, osteopenia and osteoporosis (light blue and white arrows), a marked inflammatory process is seen clearly in the synovial membrane and the adjacent articular capsule and tendon (tenosynovitis; black arrows), the predominant inflammatory cells are lymphocytes, plasma cells, macrophages and neutrophils, soft hypodermal tissue surrounding the affected joint shows aggregates of inflammatory cells (red arrows) with occasional adnexal damage and granulomatous nodular reaction (green arrows). C Methotrexate-Arthritis group showing remnants of cartilaginous and boney degenerative and necrotic changes (yellow arrows), associated with moderate inflammatory reaction in the synovial membrane (black arrows) and soft tissue (tenosynovitis; blue arrows). D Hemolymph-Arthritis group showing cartilaginous-bony regeneration with remarkable disappearance of the previous osteoporotic changes (yellow arrows), synovial membranous healing (black arrows) and resolving of most of inflammatory and/or granulomatous reactions in the surrounding soft hypodermal tissue with adnexal regeneration (blue and green arrows)
In contrast, the arthritis group exhibited severe pathological changes involving the cartilage, bone, synovium, joint capsule, and surrounding hypodermal tissue, leading to widespread tissue damage. These alterations resulted in articular cartilage degradation, subchondral bone damage, bone marrow lesions, and episodic synovitis. Articular cartilage breakdown was characterized by chondrocyte degeneration and necrosis, occasionally forming cysts. The underlying bone displayed osteolytic degeneration, osteopenia, osteoporosis, and an increased presence of inflammatory cells infiltrating the interstitial tissue. Osteoclastic activity was also evident. A pronounced inflammatory response was observed in the synovial membrane and adjacent articular capsule and tendon (tenosynovitis), with predominant infiltration by lymphocytes, plasma cells, macrophages, and neutrophils. The surrounding hypodermal tissue also exhibited inflammatory cell aggregation, occasional adnexal damage, and granulomatous nodular reactions (Fig. 6B).
The treatment groups showed varying degrees of therapeutic efficacy. The hemolymph-treated group demonstrated the most significant improvements, with notable cartilaginous and bony regeneration, resolution of osteoporotic changes, and synovial membrane healing. Most inflammatory and granulomatous reactions in the surrounding soft tissue subsided, and adnexal regeneration was observed (Fig. 6D). In contrast, the MTX-treated group exhibited only partial improvement, with residual cartilage and bone degeneration, moderate inflammatory reactions in the synovial membrane and soft tissue (tenosynovitis), and limited healing in some cartilaginous and bony structures. Partial synovial membrane recovery was noted, but moderate inflammatory cell infiltration persisted in the bone marrow and surrounding soft tissue (Fig. 6C).
OST protein expression was detected in arthritic chondrocytes, whereas both protein staining and specific mRNA expression were absent in the chondrocytes of normal adult cartilage (Fig. 7). In the control group, a few reactive osteoblasts were observed in the periosteal and perichondral tissues, as well as in bone osteoblastic cells (Fig. 7A). Positive reactivity was moderate to intense, involving both nuclear and cytoplasmic contents. In the arthritis group, a moderate number of osteoblasts and osteoclasts exhibited positive reactivity to the bone cell-stimulating marker OST (Fig. 7B).
Fig. 7
Anti-osteocalcin expression from metacarpal joint structures. A Control group showing a few reactive osteoblast cells in the peri-osteal and perichondral tissues cells. B Moderate to intense positive reactivity is seen involving both nuclear and cytoplasmic contents of a moderate number of bone osteoblastic and osteoclastic cells of the RA group (RA). C A mild to moderate number of osteoblasts and osteoclasts appears reactive to osteocalcin in methotrexate treated rats. D A few bone marrow monocytes and bone osteoblast are seen weakly expressed to osteocalcin in hemolymph treated rats. Positive cells are pointed by red arrows and negative cells are pointed by light blue arrows
In the RA + MTX group, a mild to moderate number of osteoblasts and osteoclasts showed reactivity to osteocalcin (Fig. 7C). Conversely, in the hemolymph-treated rats, only a few bone marrow monocytes and bone osteoblasts exhibited weak OST expression (Fig. 7D).
A morphometric analysis of osteocalcin-immunostained sections from the metacarpal joint structures of the control, RA, MTX-treated, and hemolymph-treated groups revealed positivity percentages of 6.75, 33.3, 16.78, and 5.81%, respectively.
Comments (0)