Our findings provide the first empirical description of the main variables associated with post-feeding dispersal of Peckia species, focused in orientation, speed, and level of aggregation. So far, quantitative studies on post-feeding mobility of Sarcophagidae species have been virtually non-existent or mostly limited to the Calliphoridae species. We infer that the data have implications for forensic entomologists and behavioural ecologists.
From a forensic entomology perspective, it is reasonable to suggest that crime scene investigations should focus sampling efforts close to the cadaver, as the estimation of the min PMI is primarily based on the age of the oldest larvae. Guidelines that suggest a wider radius (> 2.0 m) for sampling Calliphoridae larvae at the crime scene may be unrealistic, given the multidirectional movement of the insects, the difficulty of finding buried pupae, and the limitations in logistics and time associated with the work of the forensic expert (Guimarães et al. 2022).
It is difficult to contextualize our findings within the current literature because most studies focus on species of the genera of Calliphoridae Calliphora Robineau-Desvoidy, Chrysomya Robineau-Desvoidy, Cochliomyia Townsend, Lucilia Robineau-Desvoidy, and Protophormia Townsend (Andrade et al. 2002; Roux et al. 2006; Arnott and Tuner 2008; Robinson et al. 2018). The search for entomological evidence based on Sarcophagidae may therefore be spatially limited, considering that Peckia larvae travel short distances, approximately 60 cm, and can bury themselves almost immediately upon finding a suitable site, usually in less than a minute. However, despite belonging to the same genus, differences exist in the overall patterns of mobility and orientation between P. (P.) chrysostoma and P. (S.) lambens.
The experimental arena for quantifying larval behaviour provided here satisfactorily allowed the registration and interpretation of the data, which leads us to suggest its use in further studies. The artificiality of the setup cannot be ignored, we chose to use a gridded acetate base to allow for videorecording, instead of soil, but the LMA was appropriate to describe the overall pattern of mobility. There is no consensus on whether larvae move slower or faster on artificial floors (e.g., concrete, tile) compared to soil (Robinson et al. 2018). Nevertheless, the LMA was designed to allow larvae to keep moving after reaching the strip of soil, in case it “decided” the distance was insufficient for a suitable place for pupation.
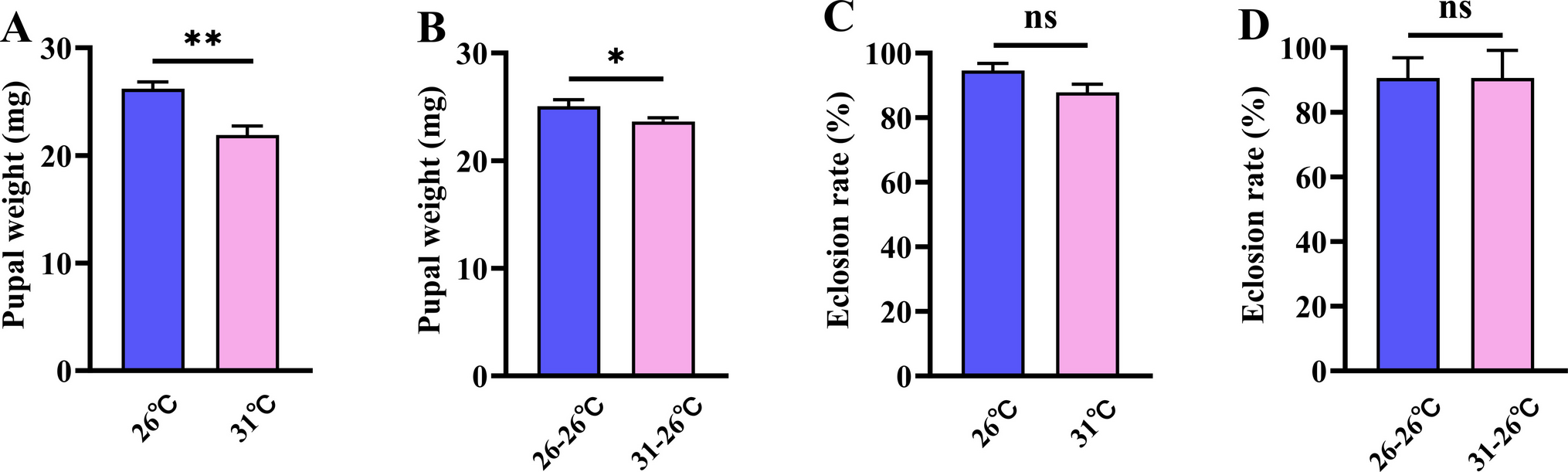
Peckia larvae moved little, but fast: it took less than a minute for P. (P.) chrysostoma to reach the soil, while P. (S.) lambens took a little longer (ca. 85 s). Despite significant differences in length and weight, P. (P.) chrysostoma can be twice as long and heavier as P. (S.) lambens, and the speed of movement was different for both species. The influence of soil humidity and undulation can be assessed in future studies, although for Lucilia illustris (Meigen) (Calliphoridae), the net distance travelled by larvae at the post-feeding stage did not differ between flat and undulating terrain (Freiburger et al. 2024).
Besides quantifying the total distances moved by each larva, our data can subsidize a model to predict the accuracy of the interaction between the observed and the predicted distances travelled by the larvae. These parameters can be used by the forensic experts to improve the rigor of sampling entomological evidence. For example, larvae found on cadavers in indoor environments may need to travel longer distances from the corpse until ideal pupation sites are readily available (Singh and Bala 2010). Thus, factors such as speed, presence or absence of obstacles, energy reserves, and body length can influence the horizontal distance larvae can travel (Gomes et al. 2006).
Roux et al. (2006), in a field study, observed that Protophormia terranovae (Robineau-Desvoidy) (Calliphoridae) exhibited irregular dispersal, similar to what was observed in this study for P. (S.) lambens. A slightly more oriented trajectory was observed for P. (P.) chrysostoma larvae, which had a higher proportion of larvae migrating to the same quadrant of the LMA. In nature, larval movement is influenced by micro-variations in factors such as soil slope, vegetation density, sunlight exposure, light direction, and humidity (Greenberg 1990; Roux et al. 2006). Biotic factors, such as the presence of predators or competing species, may also play a fundamental role in the route chosen by each larva (Faria et al. 2004; Reigada and Godoy 2005; Gomes et al. 2006).
A recent study by MacTaggart and collaborators (2025) brought interesting innovations to the methodology for describing post-larval dispersal movement, using an experimental arena known as a servosphere (Syntech TrackSphere LC-300), which allows studying insect locomotion in any direction and recording behavioural changes based on external stimuli. The authors demonstrated that the dispersal speed of larvae of the Calliphoridae species, P. terraenovae and Calliphora vicina Robineau-Desvoidy gradually decreases on solid surfaces and confirmed a negative phototropic relationship, with significantly greater displacement in the absence of light.
The LMA proposed here can incorporate several variables, such as temperature, slope, soil humidity, photophase, among others. Its simplicity, low cost, and easy assembly are additional advantages. Variations in larval mobility in relation to photophase can be quantified using the LMA, especially because the duration of light defines several aspects of the reproduction and behaviour of necrophagous flies (Ferreira et al. 2024).
The choice of the site of pupation in the vermiculite strip did not seem to be influenced by the first movers, which shows larvae wandering in a non-oriented route in the LMA. Interestingly, almost 50% of P. (P.) chrysostoma larvae pupated close to each other, aggregated in two of 25 quadrants, which does not discard the possibility of pheromone-mediated behaviour.
Regarding the time taken for the larvae until complete burial, both species exhibited a remarkably similar time (ca. 21 s), indicating that vermiculite was an adequate substrate and that the 5-cm depth was sufficient for complete burial. The complete burial protects the larvae from predation and allows a safe environment to undergo extreme modifications through the pupal stage until adulthood (Balme et al. 2012).
Burial depth must be superficial and in a soft soil, to allow breathing and the vertical ascension of the adult, so that the soil type can influence the duration of the pupal stage, with consequences for the estimation of min PMI. To illustrate that, the pupal stage of Calliphora terraenovae Macquart (Calliphoridae) was longer in clay soils compared to sandy soils (Moore et al. 2024). Pupae have been used with increasing frequency in forensic entomology as indicators of more advanced stages of the insect (Brown et al. 2015). Unfortunately, direct comparisons of our results with other flesh fly species are impossible due to the scarcity of research on Sarcophagidae.
Dry environments can increase larval mortality due to dehydration (Byrd and Tomberlin 2019; George et al. 2013), and these conditions can also directly influence larval development and pupation rates. Soil composition, particularly compaction and granulometry, can affect the larvae's ability to move and the effort required for burial. Compact soils may present a physical barrier, limiting burial depth, while looser soils facilitate digging and the pupation process (Balme et al. 2012).
However, the evidence found in this study reveals that once larvae of Peckia find an ideal location for pupation, they immediately bury themselves. For FE, finding pupae can provide information consistent with a longer period of cadaver colonization, and shallow excavation may be sufficient for sampling these species at the death scene. The age of empty puparia of Phormia regina (Meigen) (Calliphoridae) collected in the superficial soil close to a cadaver validated the estimation of min PMI in a case in Poland (Bajerlein et al. 2018). The search for pupae by forensic experts should be encouraged, as it can provide robust data for estimating the min PMI based on intrapuparial development (Dias et al. 2024).
Given the increasing relevance of pupae as entomological evidence, our findings on dispersal and burial provide insights into behavioural patterns relevant for understanding the life history of species, the decomposition dynamics in the necrobiome, and the necessity of incorporating data on insect behaviour in forensic entomology.


Comments (0)