
Remember me
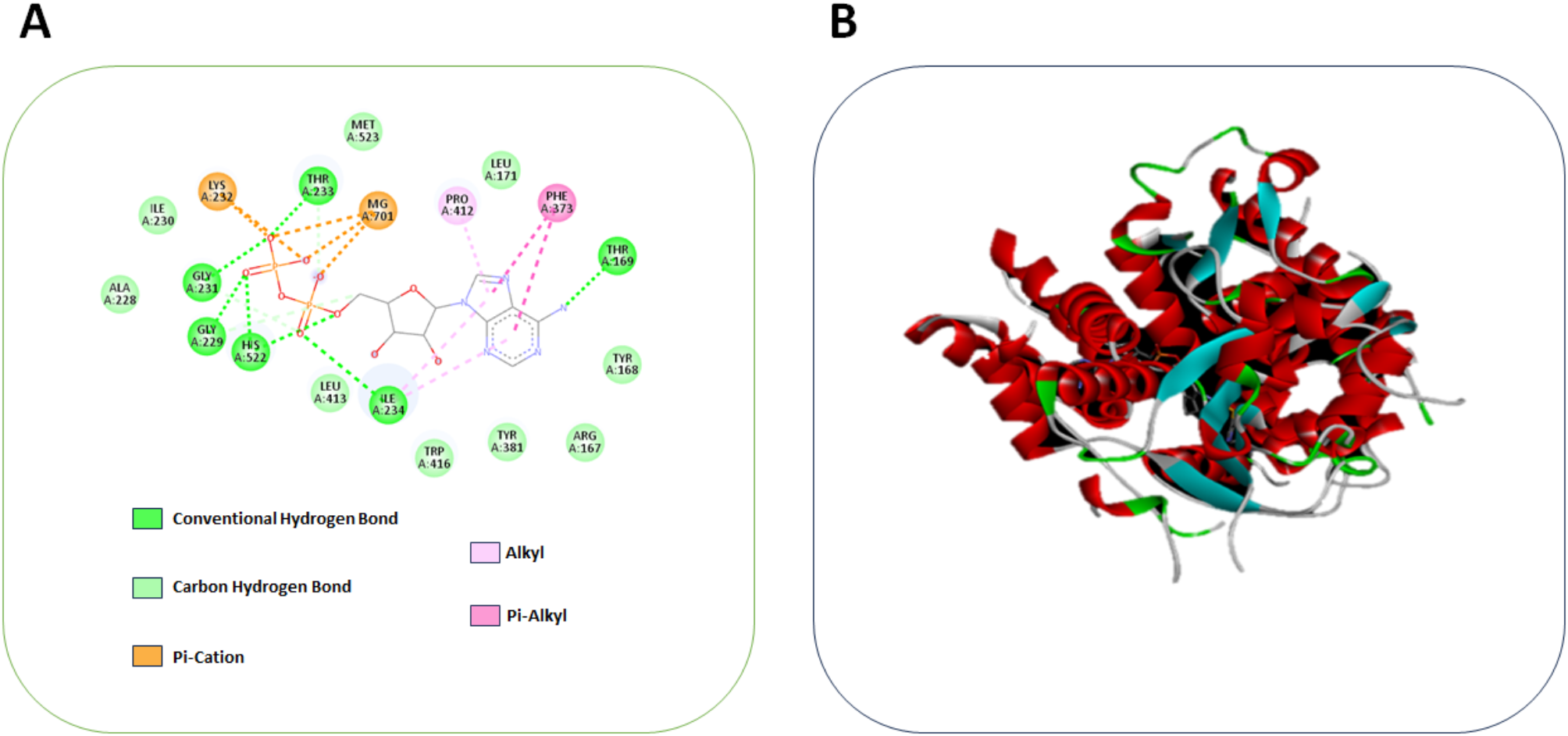






During the docking analysis, the binding sites of the protein molecule and the ligand are indicators to determine their interaction. The lowest binding affinity indicates the most significant match for bioactive compounds. In this study, the compound MV was docked against NLRP3, which was found to have a binding affinity of −7 Kcal/mol. The NLRP3 (PDB ID: 8ETR) was found to have a resolution of 3.50 Å, and they were analyzed by electron microscopy. The compound MV has better interaction with NLRP3, which includes residues GLY A:231, GLY A:229, ALA A:228, ILE A:230, LYS A:232, HIS A:522, THR A:233, MG A:701, LEU A:413, ILE A:234, MET A:523, TRP A:416, PRO A:412, LEU A:171, TYR A:381, PHE A:373, ARG A:167, THR A:169, ARG A:167, TYR A:168. 2 pi-cations, 9 van der Waals, 6 conventional hydrogen bonds, an alkyl, and pi-alkyl were obtained, as shown in Fig. 1. This suggests that a higher binding affinity can inhibit NLRP3 pathways, thereby reducing fibrosis. This analysis has selected the compound MV for further in vivo and in vitro testing. The literature revealed that NLRP9 more effectively docks with Withania somnifera to treat inflammation. In the process of developing the drug, the molecular docking method revealed structure deviation, protein compactness, stability, mobility, and hydrophobicity. These factors were essential in the drug’s development. Furthermore, they showcase the potent and efficient compounds that have the potential to interact with inflammatory proteins, thereby reducing lung inflammation. Molecular docking was one of the major methods for choosing the drug for the particular treatment that may reduce the risk of side effects and portray the drug’s mechanism of action for a better understanding of the process [34, 35].
Fig. 1
A Computational model illustrated docking of compound Malvidin against NLRP3 and B Denotes the 3D structure of MV binding to NLRP3
In vitro analysisThe in vitro analysis was used in the fields of pharmacology, toxicology, molecular biology, and genetics. They provide the molecular and cellular mechanisms, which might be difficult to observe in vivo. From the supplementary Fig. 1(A&B), the effect of MV on BEAS-2B and RLE-6TN was observed up to the concentration of 250 µg/mL. From that, it was observed that the IC50 was found to be at 100 µg/mL. So, further analysis was carried out at the concentration of 50, 100, 150 µg/mL. After this optimization, the MTT assay was carried out on BEAS-2B and RLE-6TN, induced with BLM, which were treated with different concentrations of MV (50, 100, and 150 µg/mL), and the cell viability was examined. Figure 2A illustrates the fluorescence changes of BEAS-2B and RLE-6TN cell lines at different treatment levels, demonstrating the effective cell proliferation by AO staining analysis. The BLM induced group shows less fluorescence when compared to the treated group. This proves that MV was efficiency in treating PF. Figure 2B shows that cell viability was reduced when treated with BLM; in contrast, when treated with the MV, the cell viability increased, which shows that the MV was effective in treating the PF. The BEAS-2B shows significance of p < 0.001 for the treated and induced group and has p < 0.05 for the treated group, the RLE-6TN was also found to have a similar significance. Hence, this MTT was used to prove the biocompatibility of the MV in mitigating PF. The MTT significantly reduces fibroblast proliferation and metabolic activity, showing that MV has potential as an anti-fibrotic agent. Malvidin’s antioxidant properties likely played a pivotal role in scavenging ROS and reducing oxidative stress, thereby mitigating cellular damage. Furthermore, the reduction in oxidative stress may have indirectly suppressed the activation of the NLRP3 inflammasome, a key mediator in BLM-induced fibrosis [36, 37]. One of the pathophysiological features of lung function failure (PF) is the excessive buildup of extracellular matrix elements like collagen, which causes the scarring and stiffening of the lung tissue. ROS plays a significant role in this process. Increased ROS levels can trigger fibrotic pathways through cellular damage and oxidative stress. Therefore, research has demonstrated that ROS enhances the inflammatory response by elevating PF and triggering the release of inflammatory cytokines. This implies that BLM is crucial in controlling ROS production and maintaining genomic stability during cell damage. Increased ROS production, persistent inflammation, and DNA damage brought on by BLM induction all contribute to the development of fibrosis [38, 39]. Figure 2C shows that the induction of BLM into the cell lines led to an increase in ROS levels. Conversely, when the cell lines were treated with MV, the ROS levels decreased, suggesting that the cells can regulate the oxidative damage caused by BLM. Figure 2D displays the graphical changes in the ROS level for both cell lines, which proves that the MV was effective in mitigating PF, and observed that the BEAS-2B was found to have the significance of p < 0.001 for the induced and control group and p < 0.05 for the induced and treated group. The cell RLE-6TN has the significance of p < 0.001 for the induced and control group and p < 0.05 for the induced and treated group. Malvidin and its glycosides, along with the anthocyanins in blueberries, can substantially reduce ROS levels in endothelial cells and alleviate detrimental effects by modifying the concentrations of certain critical proteins. The ROS is crucial in activating the NLRP3 inflammasome, a multiprotein complex associated with inflammation and fibrosis. Increased ROS levels work as an essential signal for NLRP3 activation, resulting in the cleavage pro-caspase-1 into its active form and the subsequent release of pro-inflammatory cytokines, including IL-1β and IL-18. The NLRP3 inflammasome is crucial in forming and progressing pulmonary fibrosis by regulating inflammatory responses, activating fibroblasts, and altering the extracellular matrix. Targeting the NLRP3 inflammasome and its subsequent effects presents a viable method for the treatment of PF [40, 41]. The wound area was analyzed using the wound-healing assay, and observed changes in cell movement at different time intervals between 0, 24, and 48 h, as shown in Fig. 3(A&B). The BLM-induced BEAS-2B and RLE-6TN cell lines have less wound recovery when treated with the MV; it has a high migration rate. Figure 3C represents the graphical representation of wound recovery; it was found to have a significance of p < 0.001 for the control and the induced group. The p < 0.05 for the induced and treated group. Inflammatory cells and pathways can directly harm lung tissue by producing elastases, cytokines, and growth factors that encourage fibrosis. This process led to fibrotic alterations and disruptions to wound-healing mechanisms. Hence, wound healing was a major factor in analyzing the PF. The in vitro analysis confirms the synthesized MV was safe and effective in mitigating PF. Further, the MV was taken to the in vivo analysis to determine the efficiency of MV in alleviating PF. Figure 3D represents the schematic representation of the wound healing assay.
Fig. 2
A Fluorescence image of BEAS-2B and RLE-6TN to determine the cell viability using AO staining, where, B The graphical analysis of cell viability where, Group I represents control, Group II for the BLM-induced group, Group III for BLM+ MV (50 µg/mL), Group IV for BLM+MV (100 µg/mL), and Group V for BLM+MV (150 µg/mL), C Indicates the fluorescence changes of ROS in both the cell lines and D Denotes the graphical changes of ROS. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
Fig. 3
A & B Indicates the wound scratch assay at various time intervals of 0 h, 24 h, and 48 h and this shows that wound healing was high when treated with MV on both BEAS-2B and RLE-6TN cell lines the images were at 4x magnification, C Graphical representation of wound healing rate at percentage, D The schematic representation of the wound healing activity in BLM-induced cell lines. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
In vivo analysisMice were utilized in the analysis; the BLM induced mice were treated with MV. The BLM plays a major role in PF; they are extensively used to test anti-fibrotic, anti-inflammatory, and antioxidant therapies. BLM is relatively inexpensive and easy to administer, providing consistent and reproducible results in animal models. They were further treated with MV and the changes have been monitored. Figure 4A represents the schematic representation of the in vivo analysis. Figure 4B shows the alterations in weight seen in mice subjected to MV therapy at various doses and those undergoing BLM-induced treatment. The BLM-induced mice model was utilized to determine the anti-fibrotic mechanism. It was observed that p < 0.001 for the treated and induced group. From this, it was observed that BLM causes significant weight loss, and when treated with MV, that significantly enhances the weight gain. Figure 4C shows the analysis of lung-wet-to-dry ratio, which was high for the induced mice model, and it was found to be decreased for the treated group with the significance of p < 0.05. Figure 4D illustrates the effect of MV on the BLM group, resulting in a significant reduction in the lung index on day 28 compared to the BLM-induced group, and it was found to have p < 0.05 when compared to the treated group.
Fig. 4
A Schematic representation of BLM-induced mice model treated with MV, B Denotes the changes in the body weight of the BLM-induced mice treated with MV, C Lung wet to dry weight ratio analysis, D Effect of Lung index, where (E-H) indicates the percentage of Macrophages, Neutrophils, Total cells, and Lymphocytes changes in BALF analysis, J Shows the schematic representation of observation of the body weight changes in BLM-induced mice treated with MV. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
Figure 4(E-H) indicates that the compound went through an analysis of BALF, revealing that the BLM-induced model exhibited elevated leukocyte levels with a significance of p < 0.001 in BALF. Treatment with MV reduces the total leukocyte count, affecting neutrophils, lymphocytes, and increasing the macrophage level. BLM therapy increases the typical proportions of neutrophils and lymphocytes and markedly lowers macrophage levels (P < 0.05). The macrophage level decreased in the BLM-induced model; however, treatment with the MV led to an increase, demonstrating the drug’s efficacy in alleviating PF. The MV has an antioxidant capacity, an anti-inflammatory effect, and a potential effect on PF. MV may inhibit fibroblast proliferation and differentiation, reducing the excessive deposition of ECM; hence, it has been proven effective in mitigating PF. MV treatment significantly reduced these fibrotic markers like decreased collagen deposition, inhibition of fibroblast activation (α-SMA), and downregulation of fibrotic mediators (TGF-β) [40, 42]. Figure 4J schematic representation of the changes in the body weight of the mice.
Examination of antioxidant and oxidative stress biomarkers in lung homogenatesThe BLM-induced PF mice model and the normal control mice have different levels of MDA, SOD, GSH, MPO, and antioxidant activities of GPx and CAT. Figure 5 (A-F) demonstrated that BLM induced higher levels of MDA and MPO with a significance of p < 0.001. Following MV treatment, there was a decrease in MDA and MPO levels (p < 0.05) between the induced and treated groups, suggesting a reduction in oxidative stress. This shows that BLM administration causes increased oxidative stress, which damages cells. When they received MV treatment following the BLM injection, the levels of SOD, GSH, CAT, and GPx in the lung tissue significantly decreased with the significance of p < 0.001. It was demonstrated that the MV treatment led to the upregulation (p < 0.05) of these antioxidant factors and prevented the decline in antioxidant enzyme activities to levels comparable to the control group. The graph revealed that MDA serves as an assay for lipid peroxidation. Compared to the healthy control group, the lung tissues of the BLM-induced mice model had higher levels of MDA, which suggests that ROS was damaging the lungs. The decrease in MDA levels indicates a significant and dose-dependent scavenging of ROS by MV at doses of 5, 10, and 15 mg/kg. The imbalance between high ROS and low antioxidant defense, which results in oxidative stress and recognized molecular initiating events, essentially causes PF [43]. Among these, NADPH, NOXs, MPO, and EMT were thought to be the main producers of ROS, which can trigger fibrogenesis in myofibroblasts and the inflammatory response. Low molecular weight endogenous antioxidant enzymes (SOD, CAT, GHx) that can boost endogenous antioxidant enzyme synthesis or remove reactive species are being suggested and developed as possible treatments for PF [44]. Antioxidant enzymes reinforce the body’s defence system against ROS. SOD can convert O•2− into H2O2, which is then catalysed by GSH-PX and CAT to produce H2O and O2. The upregulation of these genes leads to a heightened capacity of cells to withstand the effects of oxidative stress, external agents, and electrophilic compound stimuli. The decrease in antioxidant enzyme activities, such as SOD and CAT, along with an increase in NLRP3 expression prevents PF [45]. Figure 5G shows the NOx level also tends to increase in the BLM-induced model (p < 0.001). When treated with MV, the level tends to reduce significantly (p < 0.05), indicating the reduction in oxidation stress and inflammation, such as physiological and pathological processes, and that NO plays various roles in PF. NO is a signaling molecule and the amounts of nitrite and nitrate in biological fluids and tissues can reveal information about the degree of oxidative stress and NO production, which are linked to PF [39]. MV treatment significantly reduced the BLM-induced changes in lung iNOS and eNOS expressions within the BLM-induced cohort. These outcomes were attained by reducing iNOS and elevating eNOS protein, which reverted to nearly normal levels. The BLM-induced group has significantly elevated iNOS protein expression and a decrease in eNOS protein expression. The primary factor contributing to lung function failure was the synthesis of nitric oxide (NO), mediated by three isoforms of nitric oxide synthase: nNOS, located in neurons; eNOS, the constitutive endothelial type with protective vasodilatory effects; and iNOS, activated by inflammatory cells and responsible for stimulating cytokine-activated macrophages within the alveoli [40]. The constriction of the lumen of the lung artery causes a reduction of eNOS levels; however, in response to the invasion of inflammatory cells, the iNOS protein levels were increased [41].
Fig. 5
A-F The impact of administration of MV on oxidative stress and antioxidant biomarkers in lung homogenates of BLM in mice and found that MDA and MPO were found to be high in BLM-induced model when treated with MV the activity has been reduced, G NOx level which also plays the major role in PF and H indicates the HYP content the activity was significantly decreased for BLM-induced and treated group. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
The lung HYP was the major factor known as a biomarker of PF. From Fig. 5H, it was observed that the level of HYP was increased when the mice were induced with the BLM (p < 0.001), and when treated with the MV (p < 0.05), the level was reduced. As a result, measuring hydroxyproline levels provides an accurate assessment of collagen deposition and the severity of fibrosis. It is essential to comprehend the etiology of PF and assess the effectiveness of treatment approaches. By focusing on collagen synthesis and deposition, which improve lung function, the potential therapies are to lower hydroxyproline levels and lessen fibrotic changes. HYP, a significant marker of the amount of collagen in lung tissue, is commonly measured to determine the degree of fibrosis [42]. Figure 6A shows the mortality rate of the mice before and after the treatment when the BLM was induced in the mice model, which causes cell damage and leads to the death of the animal. When the BLM-induced group was treated with the MV, survival increased, which confirms the potential of MV in mitigating PF. Figure 6(B-E) shows that MV significantly reduces the overall protein concentration in the BALF of mice induced with BLM. Serum ALP, LDH, and BALF’s LDH activity were significantly (p < 0.001) higher in mice induced with BLM (group II); MV treatment further confirmed these levels by reducing the activity with a significance of (p < 0.05). BLM-induced lung injury is indicated by elevated levels of ALP and LDH, and elevated BALF-LDH activities, which indicate the degree of alveolar damage [46]. BLM instillation also resulted in a marked increase in leukocytic infiltration in peri-bronchiolar tissue, alveolar walls, and it obliterated the alveolar lumen, in addition to a marked increase in airway, vascular, and parenchymal inflammation. In terms of lung inflammation overall and alveolar wall thickness, BLM mice showed significantly higher scores than control mice [31]. After receiving MV at various concentrations, they showed a reduction in inflammation, proving the compound’s effectiveness in mitigating PF. Previous studies have demonstrated that MV may lower these inflammation gene transcription levels [47]. Figure 6(F-H) shows that the TXNIP content was measured from the lung tissue homogenate and observed as high for the BLM-induced mice model (p < 0.001) when compared to the control group. When they were treated with the MV (p < 0.05), the reduction of TXNIP in the lung tissue homogenate and the BALF content level was observed. The MV significantly reduces the lung HIFα (p < 0.05) compared to the BLM group. The TXNIP inhibiting NLRP3 inflammasome pathway could help reduce chronic inflammation and its contribution to fibrosis. Targeting TXNIP and its associated pathways offers promising therapeutic potential for mitigating pulmonary fibrosis. However, further research is needed to understand the mechanisms of TXNIP in fibrosis and to develop effective and safe TXNIP-targeted therapies [29].
Fig. 6
A Denotes the survival rate of BLM-induced mice treated with MV, where (B-E) Indicates Serum ALP, Serum LDH, BALF total protein, and LDH activity in BALF, and F-H Represents the TXNIP and HIF-1α level in lung tissue and BALF. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
Histopathological analysisThe histological examination depicts that the lungs treated with BLM cause damage with obvious lesions, cell infiltration, and tissue decomposition. The changes were analyzed by H&E staining, as shown in Fig. 7A, and found that the BLM-induced model exhibited a higher score when compared to the MV-treated group, with a significance of (p < 0.05). Figure 7B shows the MT staining, which was also the other methodology used for the confirmation of PF, which shows the level of the fibrosis and deposition of the collagen. The maximum collagen depositions occurred in BLM-induced mice model when they were treated with MV; the deposition level was reduced, hence, the compound was effectively mitigated PF (p < 0.05). Figure 7(C-D) indicates that the histological score, fibrosis score, and Ashcroft score were high in the BLM-induced model when treated with MV; the score was reduced, and hence this proves that the fibrosis level has been reduced. The MV has better activity against reducing PF. The preservation of endogenous reserves and inhibition of lipid peroxidation might also support the structural integrity of the basement membrane. Histopathological analysis confirmed the previously mentioned results [48]. MV is a potentially effective treatment option for PF because it combines anti-inflammatory, anti-fibrotic, and antioxidant properties. Furthermore, its natural origin and inclusion in common dietary sources support its favourable safety profile, and the histopathological scores confirm this analysis.
Fig. 7
A Displays the damage in mice lung tissue and was determined by H&E staining, B Indicates the changes of lung tissue by MT staining, the images were at (20X & 40X). C & D Graphical representation of the histological score, Denotes the fibrosis area percentage and, E Ashcroft’s scoring. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
qRT-PCR and Western blotting analysisNLRP3 activation regulated the induction of BLM-induced inflammatory response. In Fig. 8(A-G), IL-1β controls the signal cascade producing proinflammatory cytokines (p < 0.001) for the BLM-induced and treated group. The mRNA levels of NLRP3, ASC, and Caspase-1, as well as the protein level of NLRP3, increase in the BLM-induced mice model (p < 0.001). When treated with MV, the level has been reduced with a significance of (p < 0.05). The α-SMA, fibronectin, and collagen play significant roles in pulmonary inflammation, particularly in processes that lead to tissue remodeling and fibrosis, such as in PF. α-SMA marks the activation of myofibroblasts, which contract and deposit ECM. Fibronectin mediates cell adhesion and migration, playing a role in both inflammation and fibrosis. A primary structural protein, collagen accumulates excessively during fibrosis, leading to lung tissue stiffening and scarring. Hence, the level tended to be increased in the induced model when treated with MV, which has been reduced (P < 0.05), showing the effectiveness of MV in reducing PF. Following MV treatment, the level dropped, suggesting a reduction in inflammation associated with blocking the IL-1β signaling pathway and the NLRP3 inflammasome. Hence, the MV compound was better in treating the lung inflammation effect by inhibiting the NLRP3, thereby reducing the inflammation [49]. Figure 8(H-I) illustrates that the lung tissues of mice infused with BLM exhibited higher levels of NLRP3, and ASC protein when compared to the control groups (p < 0.001) and when treated with MV, the activity has been reduced, p < 0.05. The western blotting images obtained from the administration of MV significantly suppressed the expression of NLRP3, and ASC protein levels. This indicates that the drug is effective in mitigating PF. Numerous alarm signals, such as ATP, uric acid crystals, LPS, BLM, and different environmental irritants like asbestos, silica, and alum, can be detected by the NLRP3 in an inflammasome, which triggers an activation response. As a byproduct of NLRP3-induced macrophage activation, IL-1β is essential for lung remodeling and fibrosis because it stimulates the production of pro-fibrotic mediators and activates fibroblasts, which leads to the deposition of collagen in the lung [21, 49]. This study aims to determine whether malvidin, a naturally occurring anthocyanin, can be used to treat mice’s bleomycin-induced PF and NLRP3 inflammasome assembly. The main results point to malvidin’s ability to considerably lessen NLRP3 inflammasome activation, which mitigates the fibrotic changes in lung tissue that bleomycin induces. These results highlight the need for more research to translate these findings into clinical practice and open up new avenues for malvidin as a novel therapeutic agent for fibrotic lung diseases.
Fig. 8
A-G Represents the qRT-PCR to analyze the expression of anti-inflammatory and anti-fibrotic genes whereas H-I Graphical representation of western blotting changes in NLRP3 and ASC gene and J Western blotting images of NLRP3, ASC. Data are expressed as mean ± standard deviation, sample size (n) = 3 for each group, *P < 0.05; **P < 0.01; ***P < 0.001
Comments (0)