
Remember me
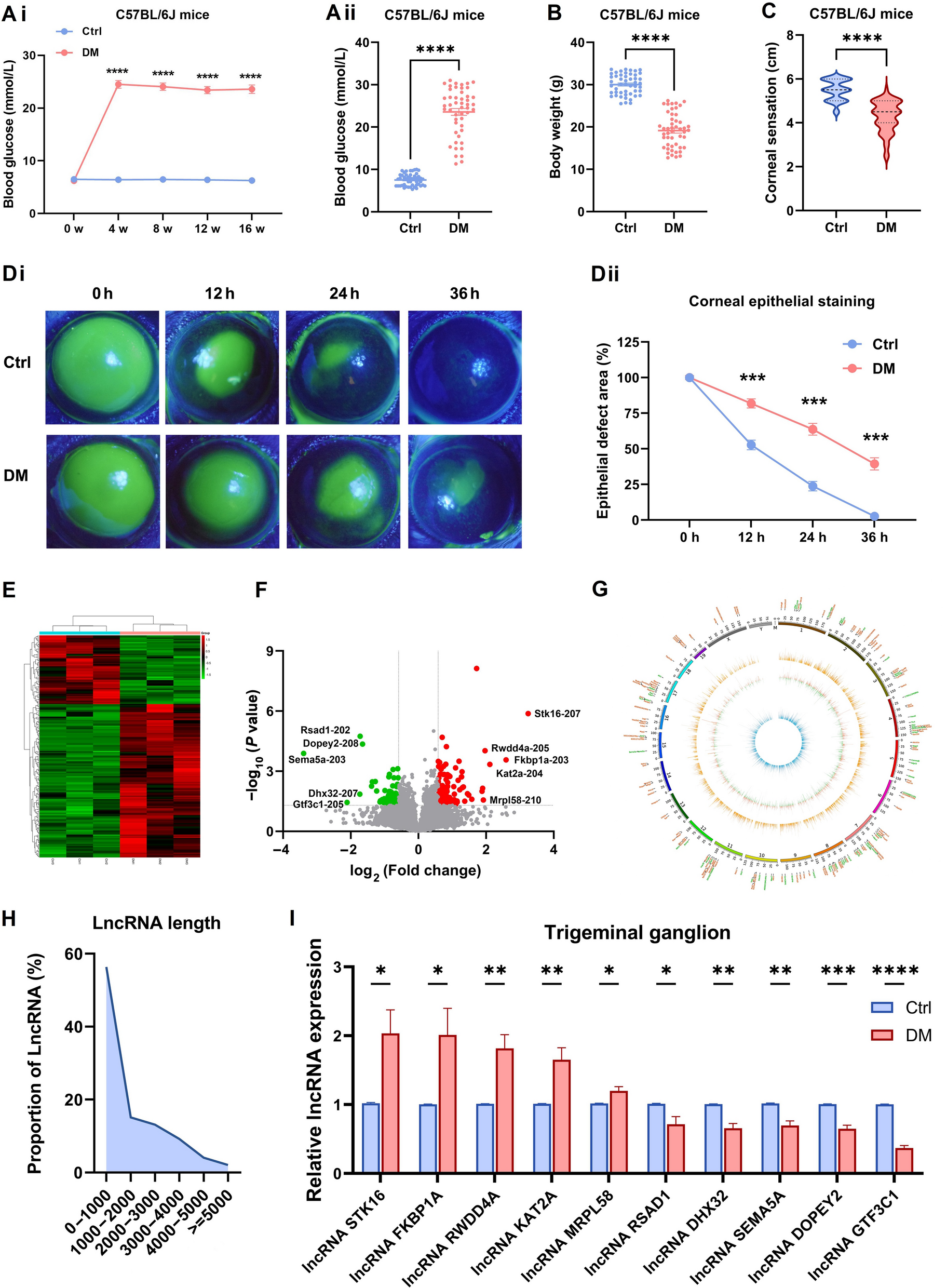




In this study, we induced type 1 diabetes in mice via the intraperitoneal injection of STZ. After induction, compared with age-matched controls, STZ-injected mice exhibited a persistent hyperglycemic state (Fig. 1A) and lost a significant amount of body weight (Fig. 1B); moreover, corneal sensitivity was significantly reduced (Fig. 1C), and corneal epithelial wound healing was significantly delayed (Fig. 1D), confirming that diabetic mice develop corneal neuropathy and delays in corneal epithelial wound healing.
Fig. 1
Differentially expressed long non-coding RNAs (DElncRNA) expression profiles and characteristics of the trigeminal ganglion (TG) tissue in diabetic mice. A: Blood glucose values of control (Ctrl) and diabetic (DM) mice. Ai: Blood glucose at 0, 4, 8, 16 weeks after intraperitoneal injection (n = 50 per group). Aii: The differences in blood glucose levels between the two groups at 16 weeks (n = 50 per group). B: Body weight of control (Ctrl) and diabetic (DM) mice at 16 weeks (n = 50). C: Corneal sensitivity of Ctrl and DM mice at 16 weeks (n = 50 per group). D: Corneas stained with fluorescein sodium at 0, 12, 24, and 36 h in Ctrl and DM mice after debridement (n = 6 per group). Di: The fluorescent staining of corneal epithelium. Dii: The percentage of epithelial defect area. E: Heatmap of the DElncRNAs in three pairs of TG tissues from Ctrl and DM mice (n = 3 per group). F: Volcano plot of DElncRNAs. Red, upregulated; Green, downregulated. G: Circos plots characterizing DElncRNAs. Outer: DElncRNAs; Orange circle: average lncRNA expression in diabetic TG; Blue: average lncRNA expression in control TG; Circle between orange and blue: the values represent the log (fold change) of diabetic TG; Orange: upregulated; Green: downregulated. H: DElncRNAs length distribution. I: Quantitative real-time polymerase chain reaction (qRT-PCR) validated the differential expression of lncRNAs in TG tissue between Ctrl and DM mice (n = 8 per group). *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001
Corneal nerve fibers are derived mainly from the TG tissue. To understand the expression profiles of lncRNAs in diabetic corneal nerves, we collected TG tissue from diabetic and control mice for RNA high-throughput sequencing analysis. A total of 194 differentially expressed lncRNAs (DElncRNA) were identified between TG tissue from diabetic (DM) mice and TG tissue from control mice (P < 0.05, fold change > 1.5), with 134 upregulated and 60 downregulated lncRNAs (Fig. 1E–F). A Circos plot was generated to visualize the expression level and chromosomal location of each lncRNA (Fig. 1G). The lengths of the lncRNA transcripts were mostly within 2000 nt (Fig. 1H). On the basis of |log2 fold change, we selected five significantly upregulated lncRNAs and five downregulated lncRNAs to verify their differential expression via qRT-PCR. The results were consistent with the sequencing trend. The most significant difference was observed for the lncRNA GTF3C1 (Fig. 1I). On the basis of the fold difference and expression abundance, we chose the lncRNA GTF3C1 for further investigation.
Identification of the lncRNA GTF3C1 as an autophagy activator in diabetic TG tissueAutophagy impairment is an important factor that delays corneal damage repair in diabetes. To study whether the lncRNA GTF3C1 is involved in the regulation of autophagy in the TG tissue, we constructed the lncRNA GTF3C1-overexpressing adenovirus Ad-GTF3C1. The subconjunctival injection of Ad-GTF3C1 successfully upregulated the expression of the lncRNA GTF3C1 in diabetic TG tissue (Fig. 2A). The experimental results revealed that LC3B protein levels were significantly reduced, while P62 protein expression was elevated in the TG tissue of DM vs. control mice, indicating that autophagy was inhibited as expected (Fig. 2B–D). Overexpression of lncRNA GTF3C1 markedly upregulated LC3B protein levels and downregulated P62 protein expression in diabetic TG tissue, suggesting that the lncRNA GTF3C1 increased the level of autophagy in diabetic TG tissue (Fig. 2B–D). In addition, transmission electron microscopy (TEM) revealed that the overexpression of the lncRNA GTF3C1 led to a higher accumulation of autophagosomes in diabetic TG tissue (Fig. 2E). These results suggest that the lncRNA GTF3C1 had an autophagy agonist function in diabetic TG tissue.
Fig. 2
Identification of the lncRNA GTF3C1 as an autophagy activator in diabetic TG tissue. A: Quantitative real-time polymerase chain reaction (qRT-PCR) validated the differential expression of lncRNA GTF3C1 in trigeminal ganglion (TG) tissue of control mice (Ctrl), diabetic mice (DM), adenovirus negative control treated diabetic mice (DM + Ad-NC), and Ad-GTF3C1 treated diabetic mice (DM + Ad-GTF3C1) (n = 3 per group) on day 5 after subconjunctival injection. B: Western blot analysis clarified autophagy proteins expression in TG tissue of Ctrl, DM, DM + Ad-NC and DM + Ad-GTF3C1 (n = 3 per group). Bi: Western blot bands of LC3B and P62 proteins. Quantified intensities of Western blot bands for LC3B (Bii) and P62 (Biii) compared with β-actin. C: Immunofluorescence analysis showed LC3B protein expression in TG tissue of each group (n = 3 per group). D: Immunofluorescence analysis showed P62 protein expression in TG tissue of each group (n = 3 per group). E: Autophagosomes (black arrows) in TG tissue of each group under a transmission electron microscope (n = 3 per group). Scale bar (including enlarged image): 50 μm. ns,not significant; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001
The lncRNA GTF3C1 promotes diabetic corneal epithelial repair and nerve regeneration by enhancing autophagyTo assess the effect of the lncRNA GTF3C1 in the repair of diabetic corneal injury, we performed subconjunctival injection of adenovirus-negative control (Ad-NC) and adenovirus containing the GTF3C1 gene (Ad-GTF3C1) in diabetic mice. Overexpression of the lncRNA GTF3C1 markedly increased corneal nerve fiber density and enhanced corneal sensitivity 5 days following corneal epithelial scraping, relative to untreated conditions (Fig. 3A, B). Moreover, the area of the corneal epithelial defect decreased significantly at 12, 24, and 26 h after scraping, suggesting that the overexpression of the lncRNA GTF3C1 promoted the healing of corneal epithelial tissues (Fig. 3C). In addition, to determine whether the lncRNA GTF3C1 affected diabetic cornea wound healing by regulating autophagy, we injected Ad-GTF3C1 into the subconjunctiva of diabetic mice concurrently with the autophagy inhibitor 3-methyladenine (3-MA). The results demonstrated that 3-MA counteracted the beneficial effects of Ad-GTF3C1 on diabetic corneal epithelial and nerve restoration (Fig. 3A–C). These results indicate that the overexpression of the lncRNA GTF3C1 promoted epithelial healing and nerve restoration in the corneas of diabetic mice by enhancing autophagy.
Fig. 3
The lncRNA GTF3C1 promotes diabetic corneal epithelial repair and nerve regeneration by enhancing autophagy. A: Corneal whole-mount staining of control mice (Ctrl), diabetic mice (DM), Ad-NC treated diabetic mice (DM + Ad-NC), Ad-GTF3C1 treated diabetic mice (DM + Ad-GTF3C1), Ad-NC and 3-methyladenine (3-MA) treated diabetic mice (DM + Ad-NC + 3-MA), Ad-GTF3C1 and 3-MA treated diabetic mice (DM + Ad-GTF3C1 + 3-MA) on day 5 after debridement (n = 6 per group). Ai: The fluorescent images of corneal nerve staining. Aii: The central corneal nerve density. Aiii: The peripheral corneal nerve density. B: Corneal sensation in each group on day 5 after debridement (n = 6 per group). C: Corneas stained with fluorescein sodium of each group at 0, 12, 24, and 36 h after debridement (n = 6 per group). Ci: Fluorescent staining of the corneal epithelium. Cii: The percentage of epithelial defect area. ns, not significant; *P < 0.05; ****P < 0.0001
The lncRNA GTF3C1 targets miR-542-3p to regulate autophagy in diabetic TG tissueTo explore the molecular mechanisms underlying the regulation of autophagy by the lncRNA GTF3C1, we first separated the nucleus and cytoplasm to determine the subcellular localization of the lncRNA GTF3C1. The results showed that more than 70% of the lncRNA GTF3C1 was expressed in the cytoplasm (Fig. 4A). lncRNAs distributed in the cytoplasm usually act as "molecular sponges" of miRNA. On the basis of previous miRNA sequencing results (GSE245193) in the TG tissue of diabetic mice, we manually identified two possible binding sites for the lncRNA GTF3C1 and miR-542-3p (Fig. 4B, Ci). To investigate the regulatory interaction between lncRNA GTF3C1 and miR-542-3p, we constructed a wild-type plasmid (GTF3C1-wt) and a mutant plasmid (GTF3C1-mut) with mutations in both binding sites (the two binding sites were spliced). The plasmids were co-transfected into human embryonic kidney 293T (HEK293T) cells with miR-542-3p mimics. The dual-luciferase reporter assay results demonstrated that miR-542-3p inhibited the luciferase activity of the wild-type lncRNA GTF3C1 reporter gene but had no significant effect on the luciferase activity of the mutant lncRNA GTF3C1 reporter gene (Fig. 4Cii). In addition, we subconjunctivally injected Ad-GTF3C1 into diabetic mice, and the expression of miR-542-3p in TG tissue was significantly lower than that in untreated diabetic mice (Fig. 4D). These results confirmed the targeting and binding relationships between lncRNA GTF3C1 and miR-542-3p.
Fig. 4
The lncRNA GTF3C1 targets miR-542-3p to regulate autophagy in diabetic trigeminal ganglion (TG) tissue. A: Relative expression of lncRNA GTF3C1 in the nucleus and cytoplasm of TG tissues from control (Ctrl) and diabetic (DM) mice detected by quantitative real-time polymerase chain reaction (qRT-PCR). B: Volcano plot of differentially expressed miRNAs. C: Dual luciferase gene reporter validated the interaction between lncRNA GTF3C1 and miR-542-3p. Ci: Potential binding site of miR-542-3p to the lncRNA GTF3C1 3' UTR and mutation site of the lncRNA GTF3C1-mut plasmid vector. Cii: Dual-luciferase gene reporter assay for relative luciferase activity after co-transfection of GTF3C1-wt and GTF3C1-mut plasmid vectors with miR-542-3p mimics or negative control in HEK293T cells (n = 3 per group). D: qRT‒PCR validated the differential expression of miR-542-3p in TG tissue of Ctrl, DM, Ad-NC treated diabetic mice (DM + Ad-NC), and Ad-GTF3C1 treated diabetic mice (DM + Ad-GTF3C1) (n = 3 per group) on day 5 after subconjunctival injection. E: Western blot analysis showing autophagy proteins expression in TG tissue of each group (n = 3 per group). Ei: Western blot bands of LC3B and P62 proteins. Quantified intensities of Western blot bands for LC3B (Eii) and P62 (Eiii) compared with β-actin. F: Immunofluorescence analysis showing LC3B protein expression in TG tissue of each group (n = 3 per group). G: Immunofluorescence analysis showing P62 protein expression in TG tissue of each group (n = 3 per group). Scale bar (including enlarged image): 50 μm. ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001
Next, we used the agomir miR-542-3p to design a rescue experiment to verify whether the lncRNA GTF3C1 modulates autophagy in diabetic TG tissue through miR-542-3p. Following subconjunctival administration of Ad-GTF3C1 in diabetic mice, subsequent injection of the miR-542-3p agomir increased miR-542-3p expression levels. At the level of autophagy proteins, the miR-542-3p agomir counteracted the upregulation of LC3B and downregulation of P62 by Ad-GTF3C1 (Fig. 4E–G), suggesting that miR-542-3p reversed the agonistic effect of lncRNA GTF3C1 on autophagy in diabetic TG tissue. The above evidence indicated that lncRNA GTF3C1 targeted miR-542-3p to enhance the autophagy level in diabetic TG tissue.
GABARAP and PTEN are potential targets of autophagy regulated by miR-542-3p in diabetic TG tissueThe function of miRNA is usually to induce the formation of RNA-induced silencing complexes (RISCs) to block target gene expression. To identify specific downstream genes regulated by miR-542-3p in autophagy, we used the TargetScan, StarBase, and miRDB databases to screen two autophagy-related target genes, PTEN and GABARAP (Fig. 5A), both of which had high binding scores. Moreover, qRT-PCR confirmed that the expression of both PTEN and GABARAP were significantly reduced in diabetic TG tissue. GABARAP, a member of the ATG8 protein family, is involved in critical cellular processes, including autophagy initiation, phagosome formation, and vesicle maturation [23]. PTEN exerts its regulatory function by negatively modulating the phosphatidylinositol 3-kinases/protein kinase B (PI3K/AKT) pathway, thereby influencing the downstream activity of mTORC1, which affects the initiation of autophagy. Through TargetScan, we obtained potential binding sites of miR-542-3p that are phylogenetically conserved in the 3' UTRs of GABARAP and PTEN and constructed wild-type and mutant plasmids of GABARAP and PTEN (Fig. 5C). The dual-luciferase reporter assay demonstrated a significant reduction in luciferase activity following the co-transfection of miR-542-3p mimics with wild-type GABARAP and PTEN plasmids. On the other hand, after the co-transfection of GABARAP and PTEN mutant plasmids and miR-542-3p mimics, luciferase activity did not change significantly (Fig. 5D). Next, we subconjunctivally injected the miR-542-3p agomir and miR-542-3p antagomir into diabetic mice to upregulate and silence miR-542-3p expression, respectively. The results indicate that the miR-542-3p agomir reduced the expression of the GABARAP and PTEN proteins in the TG tissue. The miR-542-3p antagomir upregulated the expression of GABARAP and PTEN proteins (Fig. 5D). Taken together, these demonstrate that miR-542-3p specifically interacts with GABARAP and PTEN. Hence, GABARAP and PTEN are potential targets for miR-542-3p to regulate autophagy in diabetic TG tissue.
Fig. 5
GABARAP and PTEN are potential targets of autophagy regulated by miR-542-3p in diabetic trigeminal ganglion (TG) tissue. A: The Venn diagram showed potential autophagy-related target genes of miR-542-3p were predicted in the Targetscan, StarBase, miRDB databases. B: Quantitative real-time polymerase chain reaction (qRT-PCR) validated the differential expression of GABARAP and PTEN in TG tissue of control (Ctrl) and diabetic (DM) mice (n = 3 per group). C: Potential binding site of miR-542-3p to PTEN and GABARAP (n = 3 per group). Ci: Potential binding site of miR-542-3p to the GABARAP 3' UTR and mutation site of the GABARAP-mut plasmid vector. Cii: Potential binding site of miR-542-3p to the GABARAP 3' UTR and mutation site of the GABARAP-mut plasmid vector. D: Dual-luciferase gene reporter assay for relative luciferase activity after co-transfection of plasmid vectors with miR-542-3p mimics or negative control in HEK293T cells (n = 3 per group). Di: Relative luciferase activity after co-transfection of GABARAP-WT and GABARAP-Muta plasmid vectors with miR-542-3p mimics or negative control. Dii: Relative luciferase activity after co-transfection of PTEN-WT and PTEN-Muta plasmid vectors with miR-542-3p mimics or negative control. E: Western blot analysis showing GABARAP and PTEN protein expression in TG tissue after subconjunctival injection with miR-542-3p agomir, agomir-NC, miR-542-3p antagomir, and antagomir-NC (n = 3 per group). Ei: Western blot bands of GABARAP and PTEN proteins. Eii: Quantified intensities of Western blot bands for GABARAP compared with β-actin. Eii: Quantified intensities of Western blot bands for PTEN compared with β-actin. *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001
Co-silencing of GABARAP and PTEN abolishes lncRNA GTF3C1-mediated autophagy enhancement in diabetic TG tissueTo explore whether the lncRNA GTF3C1 regulates autophagy in diabetic TG tissue through genes downstream of miR-542-3p, i.e., GABARAP and PTEN, we constructed antisense oligonucleotides (ASO) targeting GABARAP and PTEN. The Western blot confirmed that the subconjunctival injection of the GABARAP ASO successfully suppressed GABARAP protein in the TG tissue. Similarly, the PTEN ASO successfully knocked down the expression of the PTEN protein (Fig. 6A). Next, in addition to the subconjunctival injection of Ad-GTF3C1 into DM mice, we separately injected the PTEN ASO and GABARAP ASO to silence the expression of PTEN and GABARAP. Protein analyses revealed that the GABARAP ASO partially reversed the upregulated expression of the autophagy protein LC3B and the downregulated expression of P62 by the lncRNA GTF3C1 (Fig. 6B–D). The PTEN ASO suppressed autophagy by stimulating thePI3K/AKT signaling pathway, leading to elevated phosphorylated AKT and mTOR levels, which partially counteracted Ad-GTF3C1’s influence on autophagy proteins LC3B and P62 (Fig. 6B–D). In contrast, the simultaneous silencing of PTEN and GABARAP expression saw a near complete reversal of the regulatory effect Ad-GTF3C1 has on autophagy proteins LC3B and P62 (Fig. 6B–D). These results indicated that silencing GABARAP and PTEN reversed the enhancing effect of lncRNA GTF3C1 overexpression on the TG tissue in diabetic mice and that lncRNA GTF3C1 regulated the autophagy level in diabetic TG tissue largely through GABARAP and PTEN.
Fig. 6
Co-silencing of GABARAP and PTEN abolishes lncRNA GTF3C1-mediated autophagy enhancement in diabetic trigeminal ganglion (TG) tissue. A: Western blot analysis detected the expression of GABARAP and PTEN proteins in TG tissue of diabetic mice (DM), diabetic mice injected with GABARAP ASO (DM + GABAR ASO), and diabetic mice injected with PTEN ASO (n = 3 per group). Ai: Western blot bands of GABARAP and PTEN proteins. Aii: Quantified intensities of Western blot bands for GABARAP and PTEN compared with β-actin. B: Western blot analysis detected the expression of GABARAP and PTEN proteins as well as autophagy proteins and PI3K/AKT pathway protein in diabetic TG tissues with PTEN or GABARAP knockdown after overexpression of LncRNA GTF3C1 (n = 3 per group). Bi: Western blot bands of proteins. Bii: Quantified intensities of Western blot bands for p-AKT compared with AKT. Biii: Quantified intensities of Western blot bands for p-mTOR compared with mTOR. Biv: Quantified intensities of Western blot bands for GABARAP, PTEN, LC3B, and P62 compared with β-actin. C: Immunofluorescence analysis showed LC3B protein expression in diabetic TG tissues with PTEN or GABARAP knockdown after overexpression of LncRNA GTF3C1 (n = 3 per group). D: Immunofluorescence analysis showed P62 protein expression in diabetic TG tissues with PTEN or GABARAP knockdown after overexpression of LncRNA GTF3C1 (n = 3 per group). Scale bar (including enlarged image): 50 μm. ns, not significant; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001
The lncRNA GTF3C1 promotes nerve regeneration and epithelial repair in diabetic mice through PTEN and GABARAPTo assess whether PTEN and GABARAP are involved in promoting the repair of diabetic corneal damage through lncRNA GTF3C1, we subconjunctivally injected Ad-GTF3C1 to overexpress the lncRNA GTF3C1 and then subconjunctivally injected the GABARAP ASO or the PTEN ASO to inhibit GABARAP or PTEN, respectively. We also simultaneously injected the GABARAP ASO and the PTEN ASO to jointly inhibit the expression of these two proteins. After the establishment of the corneal epithelial injury model, we found that compared with those in the Ad-NC-injection group, the corneal nerve density and corneal sensitivity were significantly higher in the Ad-GTF3C1-injection diabetic group on day 5 after injury, and the corneal epithelial healing rate was also significantly higher (Fig. 7A–C). GABARAP ASO treatment attenuated the pro-regenerative effects of lncRNA GTF3C1 on both corneal epithelial wound closure and nerve regeneration, while simultaneously restoring corneal sensitivity after diabetic corneal trauma (Fig. 7A–C). Similarly, the inhibition of PTEN expression alone partially antagonized the positive effects of lncRNA GTF3C1 in promoting diabetic corneal wound healing and nerve regeneration as well as improving corneal sensitivity (Fig. 7A–C). Notably, co-inhibition of GABARAP and PTEN signaling pathways almost completely abrogated the therapeutic benefits of lncRNA GTF3C1 in diabetic corneal repair, eliminating its positive effects on both epithelial restitution and sensory nerve regeneration. On day 5 of subconjunctival injection, there was no significant difference compared with the diabetic mice in the NC group in terms of corneal central and peripheral densities as well as corneal sensitivity (Fig. 7A–C). In summary, the lncRNA GTF3C1 enhanced autophagy and promoted corneal epithelial repair and nerve regeneration in diabetic mice through GABARAP and PTEN.
Fig. 7
The lncRNA GTF3C1 promotes nerve regeneration and epithelial repair in diabetic mice (DM) through PTEN and GABARAP. A Corneal nerves whole-mount staining on day 5 of diabetic mice with PTEN or GABARAP knockdown after overexpression of LncRNA GTF3C1 (n = 6 per group). Ai Fluorescent images of corneal nerve staining. Aii The central corneal nerve density. Aiii The peripheral corneal nerve density. B Corneal sensation of each group on day 5 after debridement (n = 6 per group). C Corneas stained with fluorescein sodium of each group at 0, 12, 24, and 36 h after debridement (n = 6 per group). Ci Fluorescein-stained images of corneas. Cii Percentage of epithelial defect area. ns, not significant; *P < 0.05; **P < 0.01; ****P < 0.0001
Comments (0)