
Remember me
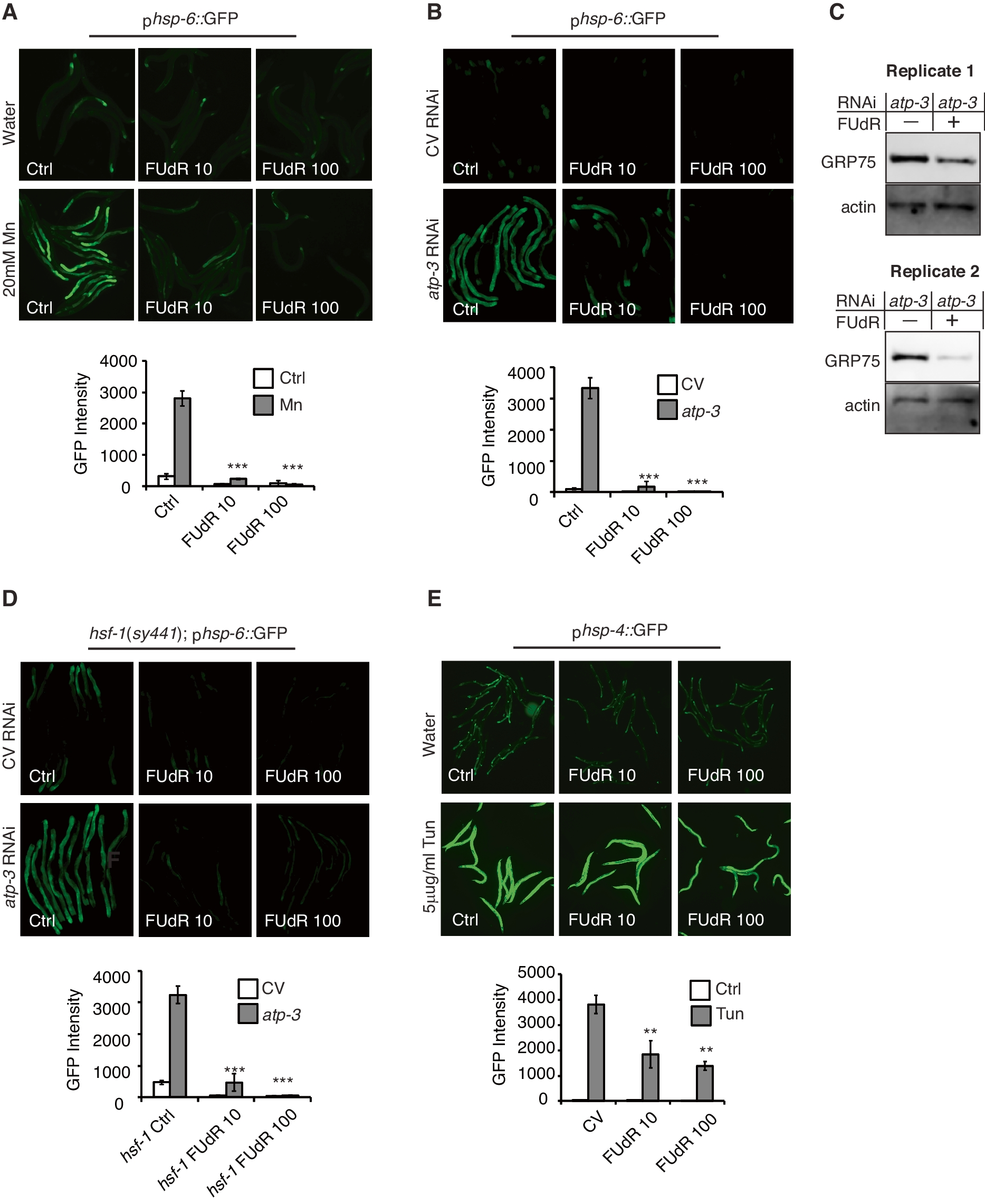
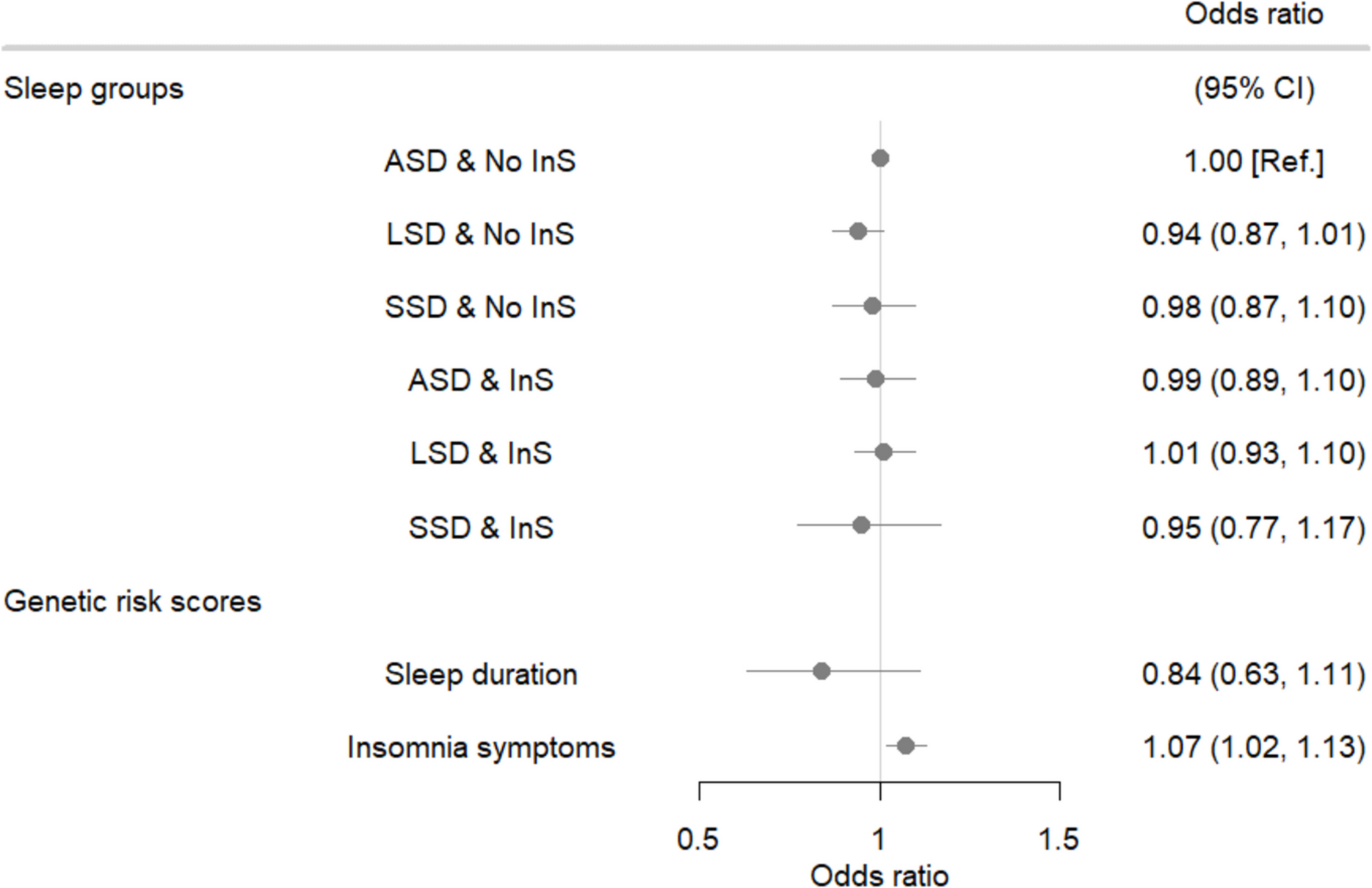

To monitor the intestinal UPRmt, we utilized the transcriptional reporter strain, phsp-6::GFP, which expresses GFP under the promoter of the mitochondrial Hsp70 molecular chaperone hsp-6 [16]. After UPRmt activation, GFP is robustly expressed throughout the intestines (Figs. 1A, B and 2A). We used two different methods to activate the UPRmt. Nematodes were treated with either 20mM manganese chloride (Mn) at the L4 stage [7] or RNAi of the F-ATP synthase subunit OSCP/atp-3 at the young adult stage [9]. We found that the concomitant addition of either 10μg/ml (FUdR 10) or 100μg/ml FUdR (FUdR 100) robustly inhibited the activation of the intestinal UPRmt by either Mn- or RNAi treatments (Fig. 1A, B) The effectiveness of FUdR as a DNA synthesis inhibitor was confirmed by the presence of unhatched embryonic eggs on plates (data not shown). To determine if FUdR treatment inhibited endogenous activation of the mitochondrial HSP-6 protein, we performed immunoblot analysis. The C. elegans HSP-6 protein is orthologous to and recognized by the mammalian GRP75/HSPA9 antibody. We found that 24 hour treatment with 100μg/ml FUdR reduced endogenous HSP-6 protein levels in nematodes that had been treated with OSCP/atp-3 RNAi (Fig. 1C), consistent with the reporter strain results. We conclude that FUdR treatment inhibits the activation of the mitochondrial UPRmt chaperone mtHsp70/HSP-6.
Given that FUdR treatment has previously been reported to activate the cytoplasmic heat shock response (HSR) [1,2,3], we sought to determine if FUdR may be affecting the UPRmt due to its activation of the HSR. We crossed a strain harboring a mutation in the master HSR transcription factor HSF1 (hsf-1(sy441)) to the UPRmt phsp-6::GFP reporter strain. We found that in the absence of deliberate UPRmt activation, hsf-1 mutants displayed a mildly elevated endogenous UPRmt compared to non-mutant controls (Fig. 1B CV Ctrl vs.1D hsf-1 CV Ctrl; **p ≤ 0.001). After RNAi of OSCP/atp-3, however, hsf-1 mutants mounted a robust UPRmt that was completely inhibited by FUdR treatments, indistinguishable from non-mutant controls (Fig. 1B, D). These findings suggest that the inhibition of the UPRmt by FUdR is independent of its effects on the HSR. To determine the impact of FUdR treatment on other stress responses, we analyzed the endoplasmic reticulum UPR (UPRER) using the transcriptional reporter strain, phsp-4::GFP, which expresses GFP under the promoter of the ER Hsp70 molecular chaperone hsp-4 [17]. We activated the UPRER via tunicamycin (Tun), a glycosylation inhibitor, in young adult nematodes and found that both low and high doses of FUdR significantly inhibited the UPRER (Fig. 1E). However, at its highest concentration, FUdR inhibited the UPRmt by eightfold or more (Fig. 1B), while it only inhibited the UPRER by 2.8-fold (Fig. 1E). We conclude that FUdR treatment is specific in its ability to robustly inhibit activation of the intestinal UPRmt.
Fig. 1
Pharmacological inhibition of the germline blocks activation of the intestinal UPRmt. A. Top panels: Photomicrographs of phsp-6::GFP reporter strain after treatment with 20mM MnCl2 in the absence or presence of 10 or 100μg/ml FUdR. Nematodes were exposed to treatments beginning at the L4 stage for 48 hours. Bottom panel: Quantification of GFP intensity in the absence or presence of 10μg/ml or 100μg/ml FUdR and 20mM MnCl2. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. ***p ≤ 0.0001 by Student’s t-test. B, D. Top panels: Photomicrographs of phsp-6::GFP reporter strain (B) or hsf-1(sy441); phsp-6::GFP reporter strain (D) after treatment with RNAi against OSCP/atp-3 in the absence or presence of 10μg/ml or 100μg/ml FUdR. Nematodes were exposed to treatments beginning at the young adult stage for 48 hours. CV, control vector RNAi. Bottom panel: Quantification of GFP intensity. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. ***p ≤ 0.0001 by Student’s t-test. C. Immunoblots from N2 nematodes after RNAi against OSCP/atp-3 in the absence or presence of 100μg/ml FUdR. RNAi and FUdR were administered for 24 h beginning at the young adult stage and then collected for analysis. Representative immunoblots are from two biological experiments. Actin was used as a loading control. E. Top panels: Photomicrographs of phsp-4::GFP reporter strain after 5μg/ml tunicamycin treatment in the absence or presence of 10μg/ml or 100μg/ml FUdR. Nematodes were exposed to tunicamycin and FUdR beginning from young adulthood for 48 hours. Bottom panel: Quantification of GFP intensity from nematodes treated with 5μg/ml tunicamycin and either 10μg/ml or 100μg/ml FUdR. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. **p ≤ 0.001 by Student’s t-test
To determine if genetic inhibition of the germline recapitulated the pharmacological effects of FUdR, we tested nematodes with a temperature-sensitive mutation in the Notch-like receptor, glp-1, which inhibits germline stem cell proliferation and results in a nearly empty gonad. Similar to FUdR-treated nematodes, young adult glp-1(e2141) mutants inhibited the intestinal UPRmt activation by sevenfold after RNAi of OSCP/atp-3 (Fig. 2A). In contrast, young adult glp-1 mutants were capable of robustly activating the UPRER as measured by the phsp-4::GFP reporter strain (Fig. 2B) and the HSR as measured by the phsp-16.2::GFP reporter strain (Fig. 2C), which expresses GFP under the promoter of the cytosolic molecular chaperone hsp-16.2 [18]. We conclude that genetic inhibition of germline stem cell proliferation specifically and robustly inhibits activation of the intestinal UPRmt.
Fig. 2
Genetic inhibition of the germline blocks activation of the intestinal UPRmt. A. Top panels: Photomicrographs of phsp-6::GFP or glp-1(e2141); phsp-6::GFP reporter strains after treatment with bacteria expressing RNAi against OSCP/atp-3. Strains were developed at 25°C to induce sterility of glp-1(e2141) mutants. At young adulthood, nematodes were shifted to RNAi plates at 20°C for 48 hours. Bottom panel: Quantification of GFP intensity. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. **p ≤ 0.001 by Student’s t-test. B. Top panels: Photomicrographs of phsp-4::GFP or glp-1(e2141); phsp-4::GFP reporter strains after treatment with 5μg/ml tunicamycin. Strains were developed at 25°C to induce sterility of glp-1(e2141) mutants. At young adulthood, nematodes were shifted to tunicamycin at 20°C for 48 hours. Bottom panel: Quantification of GFP intensity. No significant difference in GFP activation was observed between phsp-4::GFP or glp-1(e2141); phsp-4::GFP reporter strains after tunicamycin treatment. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. C. Top panels: Photomicrographs of phsp-16.2::GFP or glp-1(e2141); phsp-16.2::GFP reporter strains after heat stress. Strains were developed at 25°C to induce sterility of glp-1(e2141) mutants. At young adulthood, nematodes were heat stressed for 1 hour at 33°C, and GFP expression was analyzed approximately 5 hours later. Bottom panel: Quantification of GFP intensity. No significant difference in GFP activation was observed between phsp-4::GFP or glp-1(e2141); phsp-4::GFP reporter strains after heat stress. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments
Next, we sought to determine whether FUdR treatment or glp-1 mutations affect lifespan in the context of mitochondrial dysfunction. In the absence of FUdR, we have previously found that treatment with either Mn at the L4 stage or RNAi of OSCP/atp-3 at young adulthood (postdevelopmental treatment) leads to lifespan shortening [7, 9] (Fig. 3A, B). The same lifespan treatments in the presence of FUdR, however, completely reversed the lifespan shortening so that the nematodes’ lifespans were indistinguishable from untreated control nematodes (Fig. 3A, B). Furthermore, we and others previously identified that RNAi of OSCP/atp-3 beginning from eggs (developmental treatment) and continued throughout life leads to lifespan extension (Fig. 3C) [9, 19]. Remarkably, FUdR treatment also blocked the lifespan extension due to developmental RNAi treatment, even though FUdR treatment was not initiated until young adulthood (Fig. 3C). Next, we examined the effects of RNAi treatment of OSCP/atp-3 on glp-1 mutant lifespan. In contrast to wild type nematodes, the glp-1 mutants did not display any significant lifespan shortening after postdevelopmental treatment with OSCP/atp-3 RNAi (Fig. 3D). Thus, pharmacological or glp-1 inhibition of the germline appears to guard C. elegans from the mitochondrial perturbations in the soma, regardless of whether the perturbations shorten or extend lifespan.
Fig. 3
Pharmacological or genetic inhibition of the germline alters lifespans impacted by mitochondrial dysfunction. A. Survival curves of wild type N2s with postdevelopmental treatment of 10mM MnCl2 in the presence or absence of 10μg/ml FUdR. Nematodes were exposed to MnCl2 beginning at the L4 stage and continued throughout the entire lifespan. Representative curves were selected from three biological experiments. N2 water control (n = 189, median survival = 16 days), N2 10mM MnCl2 (n = 215, median survival = 14 days). ***p ≤ 0.0001 by Log Rank (Mantel–Cox) test. N2 water control, 10 μg/ml FUdR (n = 138, median survival = 16 days), N2 10mM MnCl2; 10μg/ml FUdR (n = 127, median survival = 16 days). p = 0.3862 by Log Rank (Mantel–Cox) test. B. Survival curves of wild type N2s with postdevelopmental treatment of RNAi against OSCP/atp-3 in the presence or absence of 10μg/ml FUdR. Nematodes were exposed to RNAi treatment beginning at the young adult stage for 48 hours and then moved to OP50 bacteria for the remainder of the lifespan. Representative curves were selected from three biological experiments. N2 CV RNAi (n = 108, median survival = 22 days), N2 OSCP/atp-3 RNAi (n = 106, median survival = 15 days). ***p ≤ 0.0001 by Log Rank (Mantel–Cox) test. N2 CV RNAi, 10μg/ml FUdR (n = 120, median survival = 19 days), N2 OSCP/atp-3 RNAi; 10μg/ml FUdR (n = 109, median survival = 20 days). p = 0.1967 by Log Rank (Mantel–Cox) test. C. Survival curves of wild type N2s treated with RNAi against OSCP/atp-3 in the presence or absence of 10μg/ml FUdR. Nematodes were exposed to RNAi treatment beginning from eggs (developmental) and for their entire lifespan. Nematodes were exposed to FUdR treatment beginning from young adulthood and for the remainder of the lifespan. Representative curves were selected from three biological experiments. N2 CV RNAi (n = 68, median survival = 15 days), N2 OSCP/atp-3 RNAi (n = 64, median survival = 26 days). ***p ≤ 0.0001 by Log Rank (Mantel–Cox) test. N2 CV RNAi, 10μg/ml FUdR (n = 67, median survival = 22 days), N2 OSCP/atp-3 RNAi; 10μg/ml FUdR (n = 64, median survival = 22 days). *p ≤ 0.05; p = 0.0466 by Log Rank (Mantel–Cox) test. D. Survival curves of glp-1(e2141) mutants with postdevelopmental treatment of RNAi against OSCP/atp-3. Strains were developed at 25°C to induce sterility of glp-1(e2141) mutants. At young adulthood, nematodes were shifted to RNAi plates at 20°C for 48 hours and then moved to OP50 bacteria for the remainder of the lifespan. Representative curves were selected from three biological experiments. glp-1(e2141) CV RNAi (n = 137, median survival = 20 days), glp-1(e2141) OSCP/atp-3 RNAi (n = 120, median survival = 23 days). p = 0.0613 by Log Rank (Mantel–Cox) test
We next sought to determine mechanistically why inhibition of germline function would impact the ability of the intestines to mount the UPRmt. Recent studies have attributed embryonic signals as critical to somatic maintenance [20, 21]. To further explore what aspect of fertility is responsible for modulating the intestinal UPRmt, we tested nematodes with a temperature-sensitive mutation in the fem-1 gene. The loss of fem-1 function leads to the complete inhibition of sperm production, resulting in a feminized (female) gonad that develops quiescent oocytes. Like FUdR-treated and glp-1 mutants nematodes, young adult fem-1(hc17) mutants did not robustly activate the intestinal UPRmt after RNAi of OSCP/atp-3 (Fig. 4A) but could activate the UPRER and the HSR (Fig. 4B, C). To determine if restoring fertility to fem-1 mutants would also restore the intestinal UPRmt activation, young adult fem-1 mutants were mated with wild type males while exposed to RNAi of OSCP/atp-3. Mating was confirmed by the restoration of egg laying in the fem-1 mutants (data not shown). Mated fem-1 mutants did activate the intestinal UPRmt after RNAi of OSCP/atp-3 during young adulthood (Fig. 4A). Our findings with mated fem-1 mutants suggest that the process of oocyte maturation, fertilization, and/or embryo deposition may modulate the intestinal UPRmt.
The FOXO/daf-16 transcription factor is known to contribute to stress resistance, pathogen resistance, and longevity in glp-1 mutants [13, 22]. Additionally, FOXO/daf-16 is required for enhanced pathogen resistance in a variety of other sterile mutants, including FEM mutants, which can upregulate FOXO/daf-16 downstream transcriptional targets [23]. We tested whether FOXO/daf-16 activity contributed to intestinal UPRmt regulation in glp-1 and fem-1 mutants. We found that introducing the daf-16(mu86) mutation completely restored the intestinal UPRmt in fem-1 mutants (Fig. 4D) and partially in the glp-1 mutants (Fig. 4E). The daf-16(mu86) mutants alone could activate the intestinal UPRmt similar to control levels (Fig. 4D, E). We conclude that FOXO/daf-16 activity contributes to the suppression of the intestinal UPRmt, though other unknown mechanisms are also involved.
Fig. 4
Late-stage reproductive signals are required for the intestinal UPRmt activation, which is partially dependent on the transcription factor FOXO/daf-16. A. Top panels: Photomicrographs of phsp-6::GFP or fem-1(hc17); phsp-6::GFP reporter strains on bacteria expressing RNAi against OSCP/atp-3. Strains were developed at 25°C to induce sterility of fem-1(hc17) mutants. At young adulthood, nematodes were shifted to RNAi plates at 20°C for 48 hours. For matings, GFP reporter strain hermaphrodites at the L4 stage were introduced to young wild type males at a 1:4 ratio for 24 hours on RNAi plates. After 24 hours, hermaphrodites were transferred to fresh RNAi plates devoid of males. Mating was confirmed by the presence of eggs for each fem-1(hc17); phsp-6::GFP hermaphrodite. Bottom panel: Quantification of GFP intensity. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. ***p ≤ 0.0001 by Student’s t-test. n. s., non-significant. B. Top panels: Photomicrographs of phsp-4::GFP or fem-1(hc17); phsp-4::GFP reporter strains after treatment with 5μg/ml tunicamycin. Strains were developed at 25°C to induce sterility of fem-1(hc17) mutants. At young adulthood, nematodes were shifted to tunicamycin at 20°C for 48 hours. Bottom panel: Quantification of GFP intensity. No significant difference in GFP activation was observed between phsp-4::GFP or fem-1(hc17); phsp-4::GFP reporter strains after tunicamycin treatment. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. C. Top panels: Photomicrographs of phsp-16.2::GFP or fem-1(hc17); phsp-16.2::GFP reporter strains after heat stress. Strains were developed at 25°C to induce sterility of fem-1(hc17) mutants. At young adulthood, nematodes were heat stressed for 1 hour at 33°C and GFP expression was analyzed approximately 5 hours later. Bottom panel: Quantification of GFP intensity. No significant difference in GFP activation was observed between phsp-4::GFP or fem-1(hc17); phsp-4::GFP reporter strains after heat stress. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. D, E. Top panels: Photomicrographs of daf-16(mu86); phsp-6::GFP or fem-1(hc17); daf-16(mu86); phsp-6::GFP (D) or glp-1(e2141); daf-16(mu86); phsp-6::GFP (E) reporter strains on bacteria expressing RNAi against OSCP/atp-3. Strains were developed at 25°C to induce sterility of fem-1(hc17) and glp-1(e2141) mutants. At young adulthood, nematodes were shifted to RNAi plates at 20°C for 48 hours. Bottom panel: Quantification of GFP intensity for either fem-1(hc17); daf-16(mu86); phsp-6::GFP or glp-1(e2141); daf-16(mu86); phsp-6::GFP reporter strains compared to controls. Data are the mean ± SEM of ≥ 45 animals combined from three biological experiments. ***p ≤ 0.0001 by Student’s t-test. *p ≤ 0.01; n. s., non-significant
Comments (0)