
Remember me
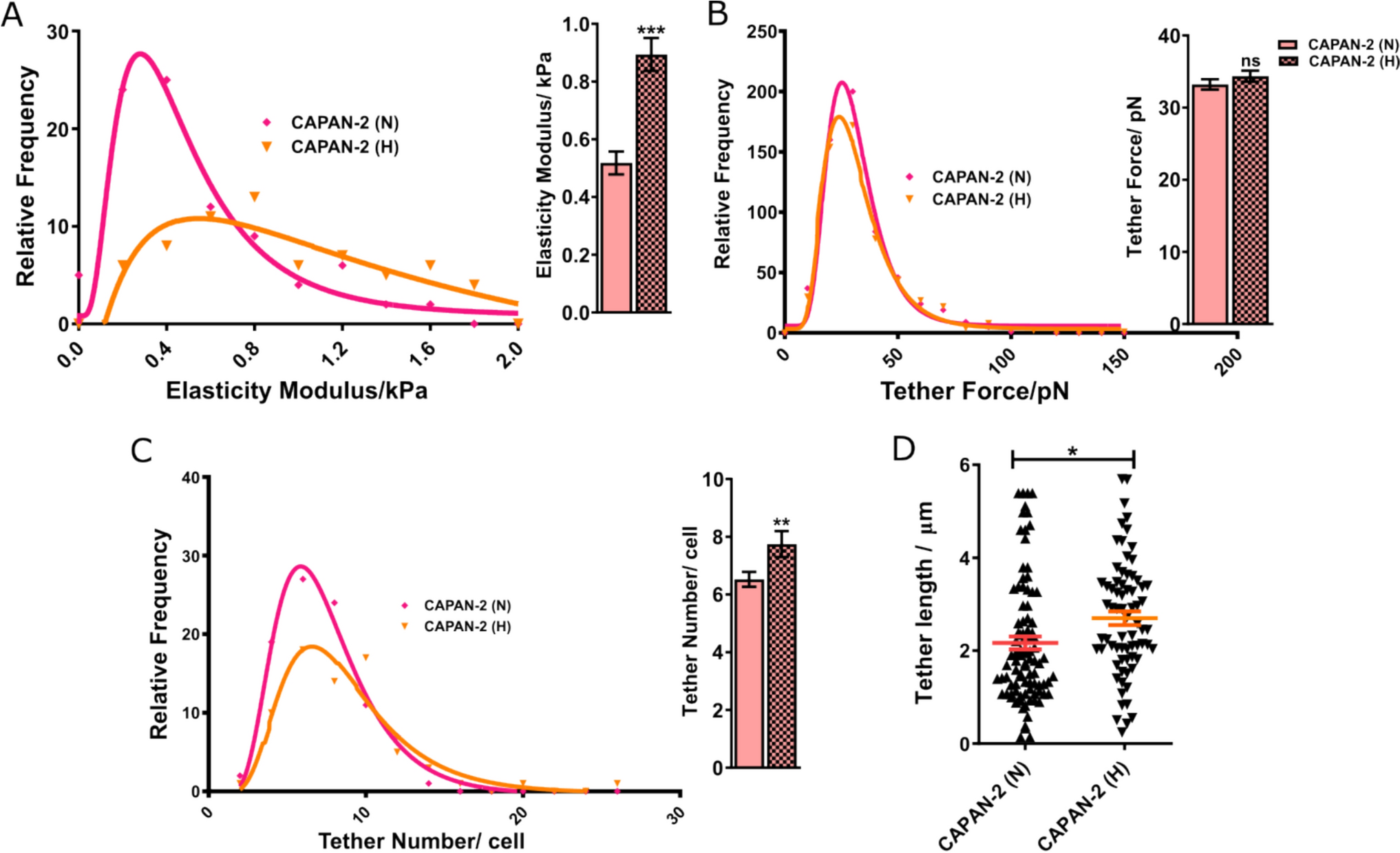
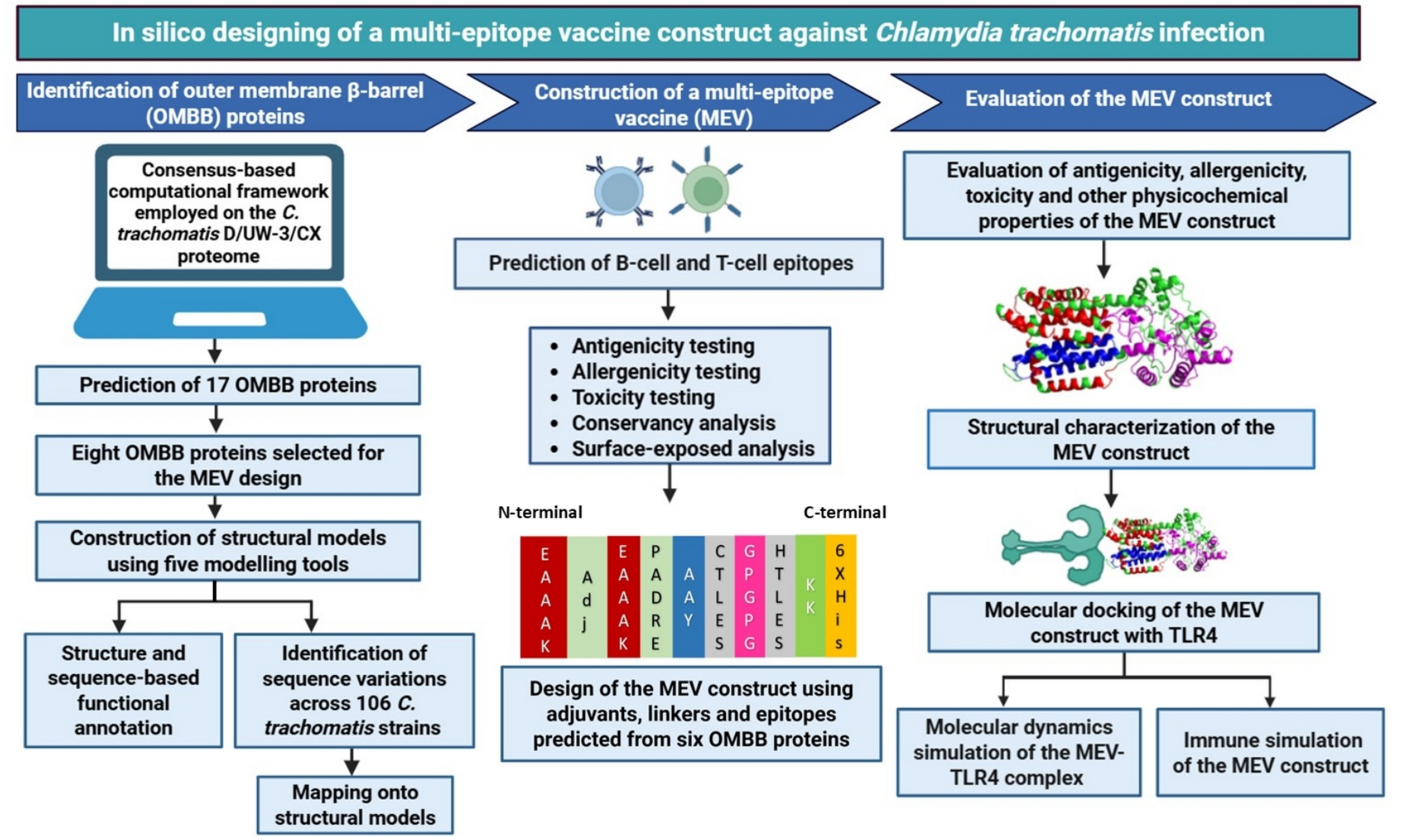
The results are presented in two sections, the analysis of the system exposed to magnetic fields with B between 0 and 10 T, and the effects observed under a field with B of 100 kT, which will be presented in the following section.
Description of the Study SystemThe system consists of 214,688 atoms, including the TIP3;1 homotetramer, the lipid bilayer and 46,255 water molecules (Fig. 1). The analysis focuses on the flow of water through the single file water channel, a region of each monomer that allows the passage of a single row of water molecules at a time. As water moves through this region, it interacts with amino acid residues in the channel and exhibits different behaviour to the rest of the water molecules. In the system, the single file water channel is located between 35 and 55 Å of the pore coordinate (z) (see Fig. 2).
Fig. 1
Description of the construction of the study system. a Homotetrameric complex consisting of TIP3;1 monomers obtained from Alphafold2. b Homotetramer immersed in a lipid bilayer fragment composed of POPC. c Initial system for simulations
Fig. 2
Representation of the single-file water channel in the study system
Water Flux Through TIP3;1 at B Between 0 and 10 TBy exposing the system under study to magnetic fields of magnitude B between 0 and 10 T, the effects on each parameter were observed and are detailed below.
Conformational Dynamics of TIP3;1The structural behavior of the TIP3;1 tetramer remained stable throughout all simulation conditions, with no significant deviations in secondary structure elements. Furthermore, the standard deviation of the pore radius profiles (Fig. 3) did not reveal structural disruption or unfolding even at the highest magnetic field intensities, confirming that the system remained within a physically realistic regime during the simulations.
Fig. 3
Standard deviation of pore radius for systems exposed to B between 0 and 10 T
Although no large-scale denaturation was detected, magnetic field exposure increased the amplitude of conformational fluctuations in loop regions and pore-lining residues. These fluctuations modulated the transient pore geometry, thereby affecting the radius and continuity of the single-file water pathway. A gradual increase in pore radius variability was observed between 2 and 6 T, suggesting that moderate magnetic intensities enhance conformational flexibility without compromising structural integrity.
Analysis of the average pore radius (r) shows consistent patterns across the systems studied (Fig. 4). A r of about 1.5 Å is observed between the 35 and 45 Å pore coordinate, with the bottleneck of the channel close to the extracellular region at about z = 50 Å. However, the r at the channel bottleneck is up to twice as large in the systems exposed at 2 T and 6 T compared to the other systems. In addition, between 35 and 45 Å of the pore coordinate, the system exposed to B of 2 T has a smaller pore radius than the other systems (1.25 Å), indicating a greater magnitude influence of the field at B of 2 T.
Fig. 4
Average pore radius resulting after exposing the study system to magnetic fields with B between 0 and 10 T
The pore radius profiles revealed two distinct responses depending on field intensity (Fig. 4). Low and moderate fields (2–6 T) induced wider and more dynamic pores, while higher intensities (> 10 T) slightly reduced the mean radius, likely due to transient conformational compaction. These changes were accompanied by variations in the hydrogen-bond network between water molecules and the pore-lining residues.
The average hydrogen-bond frequency increased under all magnetic conditions compared to the control, suggesting enhanced polarization of the water molecules and stronger transient interactions with the protein surface.
The standard deviation of the pore radius (Fig. 3) shows minimum values between 35 and 45 Å at z, gradually increasing towards the extracellular region in most treatments. This trend suggests that the maximum conformational changes occur near the bottleneck to facilitate water passage (Nieto-Giraldo et al. 2025b). In general, the systems exposed to the magnetic field have a larger standard deviation than the control, and in particular the system exposed to B of 2 T has more than twice the standard deviation of the control along the entire pore coordinate.
Thus, changes in protein conformational behaviour indicate that the magnetic field influences pore size and dynamics over the simulation time, with effects of greater intensity for systems exposed to B between 2 and 6 T, and with a smaller influence for B from 8 to 10 T. In addition, variations in pore size in the simulation can affect the patterns of protein-water intermolecular interactions in the single file water channel and the water mobility in this region.
Intermolecular InteractionsThe protein-water hydrogen bond frequency distribution (see Fig. 5) demonstrates a peak near the intra- and extracellular regions in all simulations. Systems exposed to 2 T, 4 T, or 6 T exhibit a minimum frequency between 51–54 Å, while the remaining systems demonstrate a peak in this region. Furthermore, Fig. 6 reveals that the hydrogen bonding frequency distribution in both replicates increases in systems exposed to B from 2 to 6 T, and then gradually decreases until reaching a value similar to the control system at 10 T. These variations in hydrogen bonding frequency could affect the mobility of water, since an increase in these interactions could reduce the freedom of molecular motion as suggested by Khajeh et al. (2020) and Yan et al. (2024).
Fig. 5
Frequency of protein–water hydrogen bonds along the pore coordinate for systems exposed to B between 0 and 10 T
Fig. 6
Frequency of protein–water hydrogen bonds as a function of B
Water FluxConversely, the mobility of water through the membrane fragment is also influenced by the magnetic field. This phenomenon is evident because the permeation coefficient pf increases threefold compared to the control system upon exposure to a magnetic field of 2 T to 6 T. However, the effect diminishes in systems exposed to B of 8 T and 10 T (see Fig. 7).
Fig. 7
pf coefficient as a function of B
Moreover, Fig. 8 illustrates that the magnetic field exerts a more pronounced effect on water mobility for the three subchannels located in proximity to the intracellular region. Conversely, the effect diminishes for the two subchannels that are proximate to the extracellular region. This is attributable to the presence of the channel bottleneck, which impedes water mobility and engenders a uniform and low pf across all systems.
Fig. 8
Osmotic permeability coefficient as a function of B for the five subchannels of the AQP
Although a higher number of hydrogen bonds is often associated with reduced water mobility, our results indicate that magnetic fields simultaneously increase both hydrogen bonding and osmotic permeability (pf). This apparent paradox can be explained by the interplay between hydrogen bonding and conformational dynamics. Magnetic perturbations may alter the dynamics of charged and polar side chains, affecting the local dielectric environment and facilitating larger-amplitude fluctuations in pore-lining residues. These transient widening events dominate the water transport process, compensating for the local mobility reduction that stronger hydrogen bonding would typically impose. Thus, an increased hydrogen-bond frequency can coexist with enhanced overall water flux, as transiently wider pores allow faster single-file rearrangements of hydrogen-bonded water molecules.
This interpretation is consistent with previous studies showing that field-induced conformational flexibility can modulate aquaporin conductance, as demonstrated for the electric-field–dependent behavior of the Aqy1 aquaporin (Rahimi and Lohrasebi 2020).
In summary, the effect on protein conformational dynamics, intermolecular interactions, and water mobility is higher for B magnitudes between 2 and 6 T, whereas this influence decreases for B of 8 T and 10 T. To provide an explanation for this trend, it is proposed that increasing the magnetic dose initially influences the analyzed parameters. As B increases, an energy saturation point is reached where the mentioned effects decrease in the B range of study.
Furthermore, experimental and computational studies of aquaporins have reported single-channel osmotic permeabilities in the range of 0.5–16 × 10⁻14 cm3 s⁻1, depending on the specific isoform and simulation protocol (Zhu et al. 2004; Mamonov et al. 2007; Hashido et al. 2007). The pf values obtained in the present study fall within this range, confirming that the calculated magnitudes are biophysically reasonable.
Water Flux Through AQP Exposed to Magnetic Fields with B of 100 kTExposure to the 100 kT magnetic field induced pronounced changes in the conformational dynamics of TIP3;1. The average pore profile (see Fig. 9) demonstrates a clear distinction between the control system and the system exposed to 100 kT, indicating a more pronounced channel bottleneck under the 100 kT magnetic field.
Fig. 9
Mean pore profile inside the protein channels. a Without magnetic field exposure and b exposed to a field with B of 100 kT. In c both pores are shown side by side for easy comparison, on the left the pore of the system without magnetic field exposure and on the right the one exposed to B of 100 kT
The average pore radius, as depicted in Fig. 10a, quantitatively validates the augmentation in the length of the bottleneck and the single-file water channel. This phenomenon can be elucidated by the influx of a greater amount of energy, which escalates the conformational variability, modifies the pore surface area, and alters the water displacement pathway across the membrane. This increased conformational variability is confirmed by the increase in the standard deviation of the pore radius under B of 100 kT (Fig. 10b). Exposure to B of 100 kT also significantly affected water mobility, recording a pf of (2.654 ± 0.12)·10–14 cm3 s−1, compared to (0.634 ± 0.052)·10–14 cm3 s−1 in the control system, indicating a considerable increase in water mobility across the membrane fragment.
Fig. 10
Average pore radius r (a) and standard deviation of r (b) of TIP3;1 for B of 0 T and 100 kT
Magnetic Dose Considerations in Molecular Dynamics StudiesDespite the considerations discussed above, experimental conditions in magnetobiology and those in molecular dynamics (MD) simulations differ fundamentally. Typical experiments apply B ranging from 1 mT to 1 T over periods of minutes, hours or days, whereas MD simulations operate on nanosecond timescales (Torres et al. 2018; Vashisth and Nagarajan 2009; Balakrishnan et al. 2023; Sulong et al. 2022). As shown in Table 1, magnetic field intensities above 100 kT are required in simulations to match the magnetic dose delivered in experimental setups.
Table 1 Magnetic dose as a function of B and exposure time, according to Pietruszewski, 2015 (Pietruszewski and Martínez 2015)Furthermore, even when magnetic doses are comparable, there is a considerable spatial disparity: biological systems, such as seeds, occupy volumes on the scale of cubic centimeters, while simulation boxes are constrained to nanometer-scale volumes. This spatial discrepancy precludes a direct analogy between the two approaches. Therefore, the results obtained in this study demonstrate only the existence of magnetic field effects on water transport through aquaporins, without establishing a direct correlation with experimental findings. The primary goal here is to reveal how magnetic fields influence water permeability and to elucidate the molecular mechanisms that might underlie magnetoreception. Consequently, we propose that magnetic fields can modulate cellular water flow and may act as one of the contributing factors influencing seed germination in magnetically treated seeds. However, this hypothesis cannot be definitively tested using molecular dynamics simulations alone.
Although this comparison is purely energetic and ignores macroscopic heterogeneities, it provides a rational basis for exploring stronger fields in silico to approximate the total magnetic dose delivered over experimental timescales. We explicitly stress that the 100 kT condition is exploratory and not intended to represent a biologically accessible field strength.
Perspectives and Biological ImplicationsThe impact of a magnetic field on water flow through TIP3;1 in systems exposed to an external magnetic field ranging from 0 to 10 T, and to a magnetic field with a strength of 100 kT, has been observed. The resulting changes manifest in the dynamics of protein conformations, the intermolecular interactions within the channel and the water mobility across the membrane fragment. It is noteworthy that these effects are interconnected; variations in pore size can limit or facilitate water flow by influencing the amount of hydrogen bonds in the region. In turn, these variations in intermolecular interactions have the potential to modulate the freedom of water movement.
Accordingly, one plausible mechanism of seed magnetoreception is the influence of the field on water flow through TIP3;1, which can lead to alterations in the other physiological and biological processes in which this AQP participates. Therefore, increased water mobility through the tonoplast has the potential to increase the hydrolysis of inactive forms of gibberellins stored in the vacuole, which triggers germination in a shorter time as observed experimentally. The findings suggest that magnetic fields can modulate water transport through aquaporins. Such modulation may, in turn, influence physiological processes that depend on rapid water movement, such as seed germination.
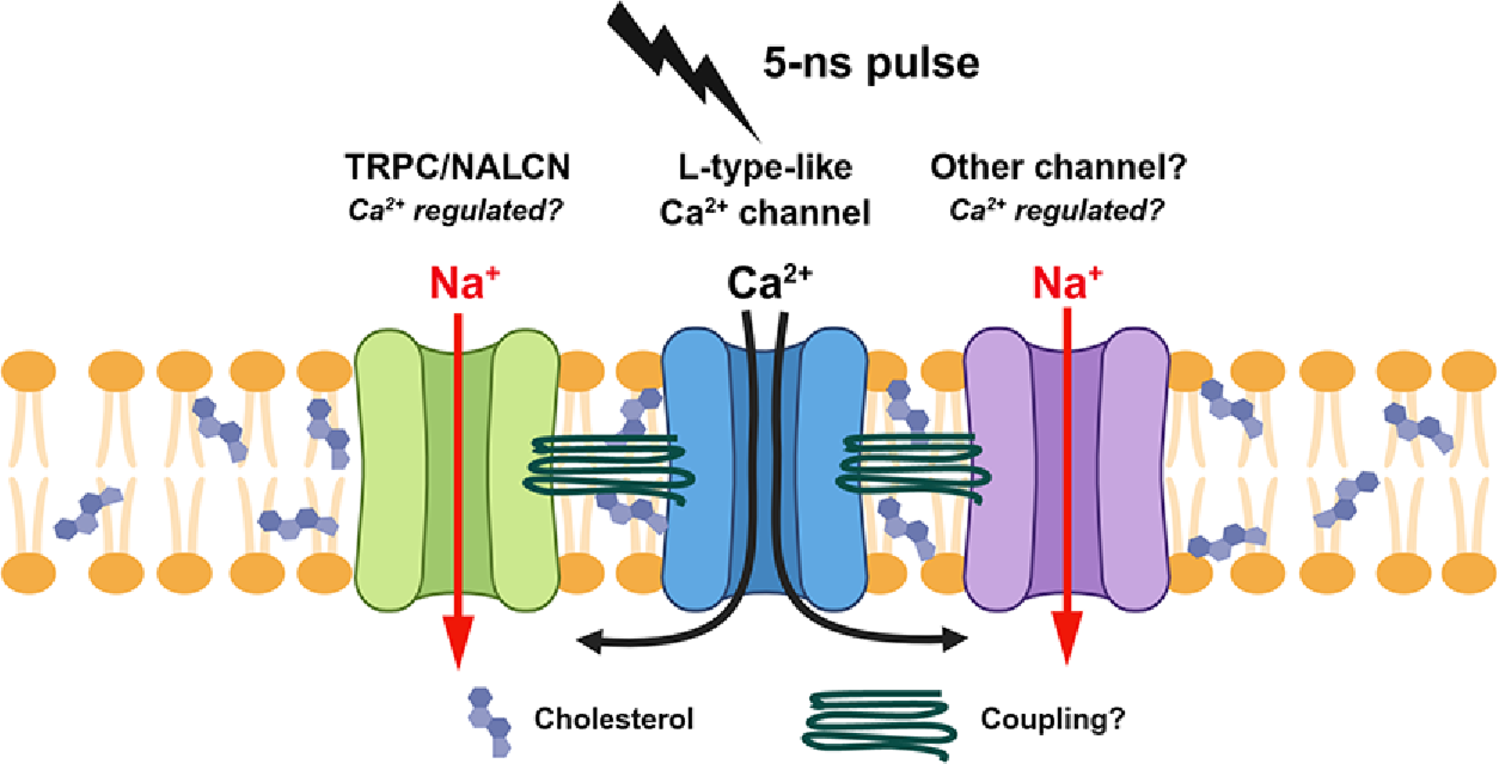
Within the broader framework of the Magnetobiology Research Group at the University of Caldas, these results provide computational support for the hypothesis that magnetically treated seeds exhibit increased GA₃ levels due to enhanced vacuolar water flow. Beyond aquaporins, magnetic fields may also affect auxiliary signaling pathways, for example, by modulating Ca2⁺ flux through the plasma membrane, which regulates Ca2⁺-dependent protein kinases responsible for aquaporin activation, or by influencing enzymatic activities such as amylases involved in seed metabolism. Further computational and experimental work will be needed to clarify these interactions.
Comments (0)