
Remember me
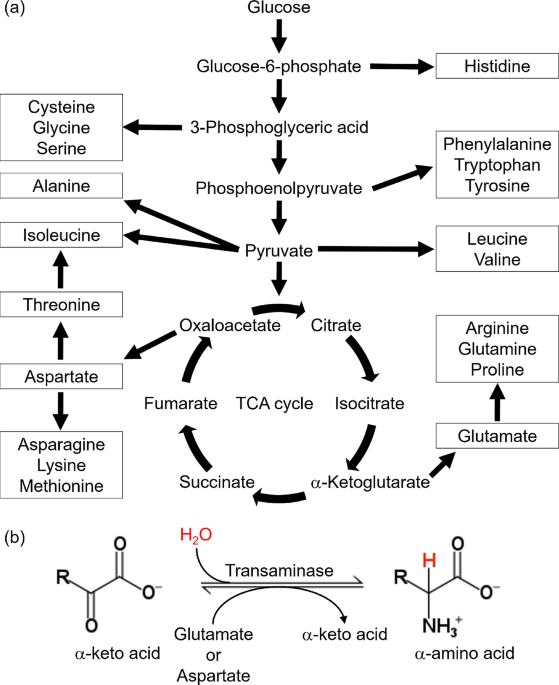

In bacterial systems, selective methyl labeling of branched-chain amino acids is typically achieved by supplementing isotopically labeled α-ketoacid precursors which are efficiently converted intracellularly into isoleucine, valine and leucine. This approach allows straightforward incorporation of 13C-methyl groups into the desired residues (Fig. 1A). In eukaryotic expression systems, only the use of α-ketoisovalerate/isocaproate precursors is feasible to label methyl groups of valine/leucine residues; however, high-molecular-weight proteins require stereospecific methyl labeling to reduce spectral overlap by restricting labeling to a single methyl group of valine and leucine. While stereospecific labeling using α-ketoacid precursor is possible, it may be affected by epimerization of stereospecifically labeled ketoisovalerate under certain conditions, resulting in residual pro-R (or pro-S) labeling. Our data indicate that epimerization of the stereospecific pro-S ketoisovalerate precursor is relatively slow in vitro, occurring on a timescale of several days under standard conditions, but still sufficient to compromise stereochemical purity upon storage or prolonged handling. To preserve stereochemical integrity, the ketoacid intermediate can either be stored at − 80 °C to significantly slow epimerization or directly converted into the corresponding L-valine, the latter strategy being adopted in this work.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Optimization of amino acid supplementation strategies to improve selective labeling and expression yield of the anti-LAMP1 antibody in CHO cells. A) Schematic overview of the synthetic pathways of isoleucine and valine. The key enzymes involved in each step are indicated in colored bubbles. B) Combined representation of anti-LAMP1 antibody production yield (left Y-axis, bars) and total amino acid concentration in the culture medium (right Y-axis, line) under different supplementation conditions (HD: High cell Density). Each mAb sample was produced only once; therefore, no error bars are shown. C) Ratio of the mean peak intensities of isoleucine to methionine residues in 2D NMR spectra, comparing cultures with or without a 16-hour growth starvation step. Ile was supplemented at 100 mg/L and Met at 1 g/L. Labeled mAb samples were generated once; therefore, error bars are not shown. D) Amino acid incorporation rates (%) of Leu, Thr and Val supplemented at 200 mg/L or 350 mg/L into the anti-LAMP1 antibody sequence, as determined by mass spectrometry
To address this, and to enable the methyl-specific labeling of isoleucine residues, we developed an efficient in vitro enzymatic synthesis protocol yielding regio- and stereoselectively 13CH₃-labeled amino acids suitable for eukaryotic expression and NMR studies of large proteins such as antibodies.
First, to produce the enzymes required for the enzymatic synthesis, recombinant ketol-acid reductoisomerase (KARI) and dihydroxyacid dehydratase (DHAD) were expressed in E. coli and purified, yielding 300 and 600 mg/L of enzyme, respectively. DHAD, a Fe-S cluster-containing enzyme, was tested under both aerobic and anaerobic (glovebox) conditions, however no significant improvement in catalytic performance was observed under anaerobic conditions during short-term assays. All subsequent reactions were therefore carried out in air with minimal handling to preserve enzyme integrity.
Using these purified enzymes, U-(2H,15N), (13CH3)δ1-Isoleucine and U-(2H,15N), (13CH3) pro-R valine were synthesized via a three-step enzymatic cascade. The first two steps, catalyzed by KARI and DHAD, were performed sequentially in the same buffer at pH 7.3. For isoleucine or valine, KARI catalyzes the rearrangement and reduction of the 2-aceto-2-hydroybutanoate precursor to its corresponding dihydroxy acid in an Mg²⁺- and NAD(P)H-dependent reaction (Dumas et al. 2001), while DHAD dehydrates the resulting dihydroxy acid to the corresponding α-keto acid (Carsten et al. 2015). The final reductive amination step, catalyzed by leucine dehydrogenase (LeuDH) at pH 9.6, yielded the amino acid with stereospecific ¹³C-methyl incorporation. Cofactor regeneration was implemented using formate dehydrogenase (FDH) to maintain NADH levels throughout the cascade.
Following enzymatic synthesis, the labeled amino acids were purified by cation exchange chromatography to remove residual precursors and by-products, preventing cytotoxicity during CHO cell culture. Overall molar yields were approximately 70–75% for δ₁-isoleucine and pro-R-valine. ¹H 1D NMR spectra confirmed the formation of Ile-δ₁-Ile and Val-pro-R with expected chemical shifts and signal intensities (Figs. S2-S3, Supplementary Data). These amino acids were then supplemented into a reconstituted chemically defined medium to produce a methyl-labeled anti-LAMP1 antibody in CHO cells.
Improved antibody expression in CHO cells through amino acid mix optimization and high-density culture for NMR applicationsThe first critical step in enabling isotopic labeling of therapeutic antibodies in mammalian cells was to establish a reliable and efficient CHO cell culture protocol for the production and purification of the anti-LAMP1 monoclonal antibody. Rich commercial media used for therapeutic production support high levels of expression but contain large amounts of unlabeled amino acids. For isotopic labeling, media containing unlabeled amino acids or supplements are unsuitable, as they dilute labeled amino acids leading to lower isotope incorporation rates. We therefore used amino acid–free media supplemented with optimized amino acid mixtures, balancing yield and labeling efficiency.
To design the amino acid mixture for isotope labeling in CHO cells, we adapted the formulation of Gossert et al. (2011), reducing it fivefold to account for the markedly lower amino acid requirements of mammalian cultures compared with insect cell systems. We tested this formulation (AAmix1 - see recipe in Table 1) in CHO cells by supplementing it into a chemically defined, amino acid–depleted medium (Fig. 1B). Under these conditions, the yield reached 25 mg/L. Doubling the concentration of the mix (2×AAmix1) did not lead to any significant improvement in yield. This suggests that, although the overall amino acid concentration was increased from approximately 3 to 6 g/L, certain amino acids may still remain limiting for optimal antibody expression in CHO cells.
These observations led us to consider whether the amino acid composition itself, rather than the total amount, might be suboptimal for CHO expression of our specific antibody. Analysis of the anti-LAMP1 sequence guided the design of a tailored amino acid mixture to better reflect the composition of the recombinant mAb. Based on this, we developed a new formulation (AAmix1_OptimAb, Table 1), in which the relative molar concentrations of individual amino acids were adjusted according to the composition of the anti-LAMP1 antibody, reflecting the relative molar fraction of each residue in its sequence (Table S2). The total amino acid content was kept to approximately 3 g/L, similar to that of AAmix1. This approach was inspired by the work of Carrillo-Cocom et al. (2015), who demonstrated that amino acid consumption in CHO cells varies according to the amino acid composition of the overexpressed protein.
Supplementing the culture medium with this customized mixture improved antibody yields to 40 mg/L, approximately a twofold increase compared to the initial AAmix1 formulation, although still lower than the yields obtained with full commercial media (80 mg/L). This suggests that the amino acids present at higher concentrations in AAmix1_OptimAb compared to AAmix1 play a particularly important role in supporting anti-LAMP1 antibody production.
Further optimization of isotope labeling in CHO cells involved timing the addition of labeled amino acids to align with the phase of active protein synthesis. During the lag and exponential growth phases, nutrients are primarily used for cell proliferation rather than protein synthesis. To avoid unnecessary isotope consumption during cell growth, we implemented a high-cell-density culture strategy, initiating cultures at 1 × 10⁷ cells/mL (the maximum viable density achievable in flasks during cell passage) instead of 1 × 106 cells/mL. Under these conditions, antibody production proceeded for 48 h before cells entered apoptosis, yielding levels (80 mg/L) comparable to those obtained with commercial rich media (Fig. 1B). This allowed recovery of 4 mg of purified anti-LAMP1 antibody from 50 mL of culture, sufficient for NMR analysis.
Enhanced methyl labeling efficiency in CHO cells through cell starvationIn our standard protocol for isotope labeling, cells were initially cultured in commercially available, unlabeled amino acid–rich medium to allow sufficient biomass accumulation. Before labeling, they were harvested by centrifugation and washed twice with amino acid-depleted medium to eliminate residual unlabeled amino acids from the extracellular environment. The cells were then resuspended in a defined labeling medium containing the desired labeled amino acids. However, despite thorough washing, intracellular pools of unlabeled amino acids, accumulated during the growth phase, can persist and be used during protein synthesis, leading to isotopic dilution (Sitarska et al. 2015). To address this, a starvation step was introduced in which cells were incubated at high density (1 × 10⁷ cells/mL) in amino acid-free medium prior to labeling. In our system, starvation durations of 4, 8, and 16 h were evaluated, and 16 h was selected based on its ability to maintain > 90% cell viability.
For labeling, U-(²H,¹⁵N), (¹³CH₃) δ¹-Isoleucine was used at 100 mg/L to remain below excess levels and enable detection of differences between the two conditions, while ¹³CH₃-methionine was added in excess (1 g/L) as an internal reference. NMR analysis of anti-LAMP1 antibody produced with and without starvation revealed a significant improvement in labeling efficiency, with the Ile/Met peak intensity ratio increasing from 0.57 to 0.76, corresponding to a 1.3-fold labeling efficiency enhancement (Fig. 1C). Importantly, antibody yield remained high (79 mg/L), confirming that this starvation step enhances isotope incorporation without compromising protein production.
To quantify the incorporation of labeled amino acids in CHO-expressed antibodies, mass spectrometry (either at the intact protein level or using peptide mapping) was performed on samples in which only one methyl-containing amino acid type was labeled at a time. This strategy allowed quantitative measurement of incorporation yield for each residue type without overlapping mass contributions. In contrast to NMR samples, non-deuterated [U-¹³C, ¹⁵N]-labeled amino acids were used to prevent mass heterogeneity arising from partial Cα-deuterium/hydrogen exchange catalyzed by cellular transaminases during protein synthesis (Sitarska et al. 2015) and to maximize the overall isotopic mass increment by uniformly labeling all carbon atoms of the considered amino acids. Incorporation rates were determined by measuring the mass shift of intact heavy and light antibody chains. For valine, threonine and leucine, supplementation with 200 mg/L of each labeled amino acids resulted in efficient incorporation with labeling levels approaching saturation around 90%. Increasing the concentration to 350 mg/L did not further improve incorporation, indicating that maximal uptake efficiency had been reached under these conditions (Fig. 1D). For methionine and isoleucine, incorporation was estimated to be greater than 84%, in agreement with the levels observed for the other residues shown in Fig. 1D. Based on these results, an intermediate concentration of 250 mg/L was selected for Methionine, Isoleucine, Valine, Leucine and Threonine, as reported in the last column of Table 1.
Improved ¹³CH₃-alanine incorporation via transaminase inhibitionAmong methyl-containing amino acids, alanine presents a specific challenge for isotopic labeling in CHO cells due to its efficient de novo synthesis from glucose via transamination, leading to significant isotopic dilution when supplemented in labeled form. For this reason, alanine was treated separately from other amino acids in our incorporation analysis. To suppress endogenous alanine biosynthesis and improve labeling efficiency, two alanine transaminase inhibitors (L-cycloserine and β-chloroalanine) were tested at various concentrations, as previously described for eukaryotic cell systems (Sitarska et al. 2015; Kofuku et al. 2018; Röβler et al. 2024) (Fig. 2A). NMR analysis of samples labeled with [¹³CH₃]-alanine revealed a 5.8-fold increase in alanine methyl peak intensity (with respect to a reference methionine signal) upon treatment with either inhibitor, regardless of compound identity or concentration, indicating a substantial reduction in unlabeled alanine incorporation (Fig. 2B). Based on these results, L-cycloserine at 10 mg/L was selected for further experiments. Incorporation efficiency was then quantified by mass spectrometry and reached 85% when 300 mg/L of labeled alanine was added to the culture medium.
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Alanine Transaminase Inhibitors Increase ¹H-¹³C NMR Signal Intensity of Alanine Residues A) Impact of alanine transaminase inhibitor type and concentration on the ¹H-¹³C NMR signal intensity of labeled alanine residues. CHO cells were cultured with U-[²H], [¹³C¹H₃]β-alanine and 1 g/L [¹³C¹H₃]ε-methionine, in the absence or presence of either β-chloroalanine (20 or 40 mg/L) or L-cycloserine (5 or 10 mg/L). Peak intensities of alanine signals were normalized to methionine signals. B) ¹H-¹³C SOFAST-methyl-TROSY spectra of U-[¹H], [¹³C¹H₃]ε-Met and U-[²H], [¹³C¹H₃]β-Ala labeled anti-LAMP1 IgG1 produced in CHO cells without (a) or with (b) 5 mg/L L-cycloserine. Methionine resonances are shown in red and alanine resonances in blue. Antibody concentrations were 40 µM (a) and 55 µM (b). Spectra were acquired at 35 °C on a 600 MHz NMR spectrometer equipped with a cryogenic probe. Each labeled mAb sample was produced only once; therefore, no error bars are shown
Selective methyl labeling enables residue-type discrimination in anti-LAMP1 antibodyUsing the established labeling protocol, five distinct anti-LAMP1 mAb samples were produced in CHO cells, each selectively labeled on a single methyl-containing amino acid: alanine, threonine, isoleucine, leucine or valine. Methionine was included in all samples as a common reference label. For isoleucine, leucine and valine, labeling was regio- or stereo-specific (Ile-δ1, Leu-pro-S, Val-pro-R).
¹H-¹³C SOFAST-methyl-TROSY spectra were acquired for each sample, revealing distinct and well-resolved cross-peaks for alanine (Fig. 3a), isoleucine (Fig. 3b), leucine (Fig. 3c) and methionine residues (Fig. 3 red peaks on the spectra). While some spectral overlap was observed for threonine (Fig. 3d) and valine (Fig. 3e) methyl groups, the overall resolution and signal-to-noise ratios were sufficient to support unambiguous residue-type classification as approximately 210 resolved peaks of the 218 expected signals (over 96%) can be seen on the spectra (Fig. 3f). These results confirm that the method enables detailed NMR analysis of CHO-expressed antibodies and provides a strong foundation for subsequent methyl resonance assignments.
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.2D ¹H-¹³C SOFAST NMR spectra in the methyl region of the anti-LAMP1 antibody produced in CHO cells and labeled with [¹³CH₃]-methionine and: a) [U-²H, ¹³CH₃]-alanine, b) [U-²H, ¹³CH₃-δ1]-isoleucine, c) [U-²H, ¹³CH₃-proS]-leucine, d) [U-²H, ¹³CH₃]-threonine, e) [U-²H, ¹³CH₃-proR]-valine. Experiments were acquired at 35 °C on spectrometers equipped with cryogenic probes operating at ¹H frequencies of 850 MHz (a, d), 950 MHz (c, e), and 600 MHz (b). To better visualize peaks, multiple zoom levels were applied for spectra (a), (d) and (e) (red and black rectangles). Methyl peaks corresponding to methionine residues are shown in red on each spectrum. (f) Number of peaks counted on the spectra compared to number of peaks expected from the Anti-Lamp1 mAb sequence for each methyl bearing amino acids
Structural integrity of methyl-labeled antibodies confirmed by NMR spectral comparisonValidation of the labeling strategy was achieved by overlaying 2D 1H-13C methyl SOFAST-methyl-TROSY NMR spectra of antibodies selectively labeled on each methyl-containing amino acid (Ile-δ1, Leu pro-S, Val pro-R, Met, Thr, and Ala) with the reference spectrum of the unlabeled antibody at natural isotopic abundance. The reference spectrum was recorded at 35 °C for 70 h on a Bruker 850 MHz spectrometer equipped with a cryogenically cooled probe, using a 0.25 mM sample. By comparison, the SOFAST-methyl-TROSY spectra of the selectively labeled antibodies were acquired with experimental times ranging from 1 to 4 h, depending on the sample, as detailed in Supplementary Table S1. As shown in Fig. S4, methyl resonances from labeled samples closely aligned with their natural abundance counterparts, indicating that isotope incorporation did not alter the mAb 3D structure. This spectral overlap demonstrates the structural integrity of the antibody under labeling conditions and confirms the reliability of the developed labeling protocol. Importantly, because valine, leucine and isoleucine residues were labeled on only one of their two methyl groups, the number of observable methyl signals in the labeled spectra was reduced compared to the natural abundance reference. This reduction led to decreased spectral crowding, thereby improving spectrum quality.
Resonance matching between cell-free produced anti-LAMP1 Fab and Fc and CHO-derived mAb supports assignment transferabilityWhile the results described above represent a major advance, demonstrating for the first time successful labeling of all six methyl-containing amino acids in a CHO-expressed antibody, they also highlight a key limitation. Without assignment of each NMR signal to specific atoms in the antibody’s 3D structure, it remains impossible to directly link spectral differences to functional or conformational changes. Such assignments would unlock the full structural potential of 2D NMR spectra, which remain underexploited in fingerprinting studies of mAbs. However, assigning methyl resonances in a 150 kDa mAb typically requires uniformly deuterated samples, which remains a major bottleneck for atomic-resolution NMR of full-length antibodies due to the toxicity of D₂O in eukaryotic cell cultures (Siegel et al. 1960).
To advance towards the assignment of methyl resonances in the full-length anti-LAMP1 monoclonal antibody expressed in CHO cells, a divide-and-conquer approach was adopted. As part of this strategy, the 13CH3 methyl labeled Fab fragment of the anti-LAMP1 antibody was previously produced in-house using a cell-free system, allowing the assignment of approximately 84% of its methyl resonances (Henot et al. 2026). A similar strategy was applied to the Fc fragment, leading to the assignment of 94% of its methyl groups (Vibert et al. 2026).
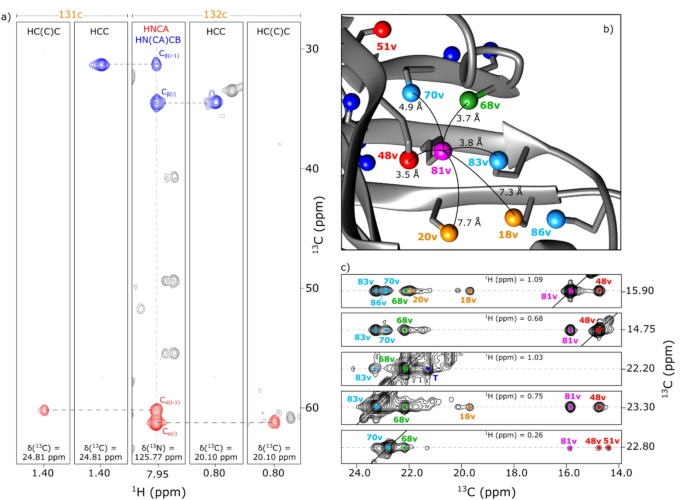
The 2D 1H–13C SOFAST-methyl-TROSY spectra of this cell-free expressed Fab fragment were then overlaid with those of the full antibody produced in CHO cells to assess structural consistency and enable resonance transfer. As shown in Figure S5, the methyl signals from alanine, methionine, leucine-pro-S, isoleucine-δ1, threonine and valine-pro-R exhibited perfect spectral overlap between the Fab and the full-length mAb, confirming that the structure of the cell-free produced isolated fragment was preserved and allowing the transfer of assigned resonances (Fig. 4). For the Fc, the assignment transfer was less straightforward. Using the previously published results of Vibert et al. (2026), the methyl resonances of the non-glycosylated Fc fragment produced in the cell-free system were assigned, and this assignment was then transferred to the glycosylated Fc fragment obtained from the CHO-expressed mAb, before finally transferring the assignments to the full-length antibody.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Assigned 2D 1H-13C SOFAST-methyl-TROSY spectra of the anti-LAMP1 mAb produced in CHO cells and specifically labeled on methyl groups. (a) mAb labeled on methionines-ε and alanines-β, zoom on alanines-β region. (b) mAb labeled on methionines-ε and isoleucines-δ1, zoom on isoleucines-δ1 region. (c) mAb labeled on methionines-ε and leucines-δ-pro-S, zoom on leucines-δ region. (d) mAb labeled on methionines-ε and valines-γ-pro-R, zoom on methionines-ε region. (e) mAb labeled on methionines-ε and threonines-γ, zoom on threonines-γ region. (f) mAb labeled on methionines-ε and valines-γ-pro-R, zoom on valines-γ region. Each assigned signal is annotated with the corresponding residue number: blue for the Fc fragment, red for the Fab fragment, bold for the heavy chain, and normal for the light chain. In panels e and f, contour levels were adjusted for some peaks, which are indicated with grey rectangles
This enabled successful assignment transfer for 97% of alanines, 100% of isoleucines-δ1, 96% of leucines-δ-Pro-S, 100% of methionines, 50% of threonines and 93% of valines-γ-Pro-R, resulting in an overall assignment of 84% of methyl containing residues in the mAb spectrum (table S3). The lower percentage of assigned threonines was due to their location in the Fab region, where most threonines were solvent-exposed and either exhibited mainly intermethyl NOE cross peaks with other unassigned threonines or showed no intermethyl NOE connectivity (Henot et al. 2026). Excluding threonines, the assignment transfer exceeds 95% of methyl-containing residues. This validates the use of cell-free expressed Fab and Fc as reliable surrogates for methyl resonance assignment in a full-length CHO-expressed antibodies.
Comments (0)