
Remember me








Mucosal-associated Invariant T (MAIT) cells are an underexplored specialized subset of T cells that play a crucial role in mucosal immune responses, serving as a bridge between innate and adaptive immunity. Their unique biology and function are defined by the expression of a semi-invariant T cell receptor (TCR), composed of an α-chain variable region (Vα) 7.2 (TRAV1-2) and an α-chain joining region (Jα) 33 (TRAJ33) paired with a limited set of beta chains (TRBV6, TRBV13, TRBV19, or TRBV20), enabling MAIT cells to recognize microbial metabolites from the riboflavin biosynthesis pathway presented by the monomorphic MHC class I-related molecule, MHC-related protein 1 (MR1) [1,2,3,4]. The semi-invariant TCR of MAIT cells and their restricting element, MR1, are highly conserved across mammals, underscoring their non-redundant roles tied to antigenic specificity, especially in mucosal and blood where they represent a substantial proportion of T cells [4, 5].
The ability of MAIT cells to recognize a broad range of microbes, coupled with their abundance and rapid, innate-like effector functions, suggests that they play a key role in human immunity. As mentioned, the primary antigens recognized by MAIT cells are metabolic adducts from the riboflavin biosynthesis pathway. These metabolites are produced by many pathogenic bacteria, as well as members of the intestinal microbiota. Interestingly, these microbial ligands can cross epithelial barriers and circulate throughout the body. This indicates that MAIT cells may play a role in real-time immune surveillance of microbiota dysbiosis, even across intact epithelial barriers [5]. Beyond their microbial recognition, MAIT cells exhibit functional plasticity, producing diverse cytokines and cytotoxic molecules, such as IL-17, IL-22, IFN-γ, and Granzyme B, which enable them to modulate local immune responses, promote tissue repair, and maintain epithelial integrity [6]. However, their roles extend beyond infectious diseases: emerging evidence highlights their involvement in non-communicable conditions such as cancer, autoimmune diseases, and chronic inflammatory disorders [7,8,9]. These diverse roles in protection and pathology underscore the versatility of MAIT cells in immune regulation and their potential as therapeutic targets.
In this review, we examine multiple aspects of MAIT cells, from their biology and development to their roles in diseases and therapies. We discuss the phenotypic and functional diversity of MAIT cells, their interactions with the microbiota, and their contributions to immune surveillance and tissue homeostasis. Additionally, we explore the emerging therapeutic potential of engineered MAIT cells, particularly in cancer immunotherapy, highlighting recent advances and identifying critical areas for future research.
Biology and development of human MAIT cellsMAIT cells are unique lymphocytes whose development begins in the thymus shortly after birth, where positive selection is mediated by MR1-expressing double-positive cortical thymocytes [3]. Emerging evidence suggests that this process may also involve selection by self-antigens presented in the thymus, MR1 in the absence of ligand binding, or microbial-derived antigens from commensal flora [10].
The selection mechanism of MAIT cells critically depends on signaling via the MAIT TCR alongside engagement of signaling lymphocytic activation molecule-associated protein on cortical thymocytes. Following positive selection, MAIT cell precursors progress through distinct developmental stages, transitioning from CD24⁺CD44⁻ to CD24⁻CD44⁻ through downregulation of CD24 and upregulation of transcriptional regulators such as KLF2, CD62L, and S1PR1, mirroring the differentiation trajectory of conventional T cells. This stage marks the initial expression of the lineage-defining transcription factor promyelocytic leukemia zinc finger (PLZF) [3, 11, 12]. Terminal differentiation yields CD24⁻CD44⁺ cells that further diversify into functional subsets characterized by the expression of T-bet (MAIT1 subset) or RORγt (MAIT17 subset). These MAIT cell subsets exhibit distinct functional profiles and tissue tropism: MAIT1 cells preferentially localize to the spleen, lymph nodes, and liver, where they respond robustly to danger signals by expressing natural killer (NK) cell-associated receptors and producing cytotoxic molecules, including interferon-γ (IFN-γ). In contrast, MAIT17 cells are enriched in barrier tissues such as the lung, skin, and gut, where they predominantly secrete IL-17 and IL-22, mediators implicated in tissue repair and homeostasis [11, 13].
MAIT cell development also depends on a range of regulatory factors, including cytokine signaling and thymic selection. During positive selection, MR1 presents antigens, such as vitamin B derivatives, driving progression through three distinct developmental stages. Stage 1 MAIT cells are characterized by CD3⁺MR1-tet⁺CD27⁻CD161⁻IL-18R⁻ markers. In stage 2, CD27 expression increases, transitioning the cells to a CD3⁺MR1-tet⁺CD27⁺CD161⁻ phenotype. Finally, stage 3 cells acquire high IL-18R expression, exhibiting the signature CD3⁺MR1-tet⁺CD27⁺/⁻CD161⁺IL-18R⁺ profile. Importantly, the use of MR1 tetramers is critical for accurately detecting MAIT cells, as monoclonal antibodies may misidentify Vα7.2⁺ non-MAIT cells or exclude earlier-stage MAIT cells. Notably, MAIT cells continue to mature and develop distinct gene expression profiles after exiting the thymus, influenced by their tissue localization. For instance, MAIT cells in the liver, lungs, and gut exhibit distinct functional profiles tailored to their respective environments. This ongoing maturation supports their functional specialization in immune surveillance and barrier tissue maintenance.
In summary, advances in understanding MAIT cell biology have clarified their developmental trajectory and highlighted the distinct characteristics of each stage. This progress has also emphasized the need for precise detection methods to accurately distinguish MAIT cells from non-MAIT T cells, further advancing research into their roles in health and disease.
Phenotypic and functional characterization of human MAIT cellsBeyond developmental markers, additional characterization markers are essential for identifying and isolating MAIT cells, enabling subset distinction, activation state tracking, and exploration of functional roles in health and disease (Table 1). MAIT cells are broadly classified into circulating and mucosal subsets, each defined by distinct steady-state and activation phenotypes, which facilitate their differentiation and characterization. In humans, circulating MAIT cells within the peripheral blood exhibit a homogeneous expression of IL-18Rα, CD161, and CD26, which facilitates their identification via cytometry [4, 14]. These cells display an effector memory phenotype (CD45RO+, CD27+, CCR7−, CD44high, CD62Llow) and express an array of integrins and tissue-homing chemokine receptors (CCR5high, CCR6high, CXCR6high, and CCR9int), indicative of their capacity for migration toward sites of local inflammation [2, 4, 6, 14,15,16,17,18,19,20,21]. In contrast, mucosal MAIT cells are distinguished by the co-expression of tissue-resident markers CD69 and CD103. Although they share the effector memory phenotype observed in circulating MAIT cells, mucosal subsets exhibit higher levels of cutaneous lymphocyte-associated antigen [22, 23].
Table 1 Phenotypic and functional markers of human MAIT cellsMAIT cells also exhibit population heterogeneity based on CD4/CD8 expression. Circulating MAIT cells predominantly consist of CD8+ (~ 70%) and double-negative (~ 15%) subsets, with a low frequency of CD4+ MAIT cells (~ 5%) [24]. In contrast, the double-negative subset is enriched within mucosal tissues, comprising ~ 40–50% of the total MAIT cell population [24]. Functionally, CD8+ MAIT cells demonstrate superior pro-inflammatory potential, characterized by higher expression of CD16 and NKG2D, elevated production of TNF-α and IFN-γ, and increased cytotoxic capacity, as evidenced by Granzyme B content and CD107A expression upon stimulation [25].
Activation mechanisms and functional regulation of human MAIT cellsFunctional markersMAIT cells can be activated through both TCR-dependent and TCR-independent mechanisms. In the TCR-dependent pathway, microbial metabolites presented by MR1 molecules, in conjunction with co-stimulatory signals from cytokines or toll-like receptors (TLRs), initiate MAIT cell activation. This enables MAIT cells to enhance local immune responses, including promoting macrophage-mediated phagocytosis and recruiting immune cells to sites of inflammation [24]. Additionally, MR1-expressing cancer cells can serve as direct targets for MAIT cells, suggesting a potential role in tumor surveillance. In TCR-independent pathways, MAIT cells can also be activated by pro-inflammatory cytokines such as IL-12, IL-18, and IFN-γ. This implies their involvement in antiviral defense, as studies have demonstrated the activation of CD161+Vα7.2+ MAIT cells in human peripheral blood during clinical infections with viruses such as dengue, hepatitis C, and influenza A [26]. The ability of MAIT cells to respond to viral infections without TCR engagement suggests a broader role in host defense beyond bacterial recognition.
When activated, MAIT cells upregulate specific surface markers and cytokines indicative of their activation. Notably, markers such as CD69 and CD25 are commonly associated with MAIT cell activation. CD69, a membrane-bounded C-type lectin receptor, is an early activation marker expressed on T cells, including MAIT cells [27]. Increased surface expression of CD69 has been reported in inflammation in autoimmune diseases. In particular, patients with ulcerative colitis exhibit a higher frequency of CD69+ MAIT cells compared to healthy controls, underscoring its role as a marker of immune activation in diseases [28]. Additionally, in psoriasis, elevated CD69 expression on circulating MAIT cells in patient peripheral blood further supports its utility as a key indicator of their activation [29]. Another important activation marker is CD25, which is also known as the IL-2 receptor alpha chain. Experimental findings have demonstrated that CD25 is upregulated following viral stimulation, serving as a reliable indicator of immune activation [30, 31]. In addition, the degranulation marker CD107a is also upregulated. The activation of MAIT cells not only involves surface marker upregulation but is also characterized by the production of various cytokines and cytotoxic molecules, which differ depending on the pathway involved. In TCR-dependent activation, rapid immune response is characterized by increased expression of RORγt and the production of cytokines such as IL-17A, TNF, and CSF2, indicative of a Tc17-like phenotype [32, 33]. In contrast, TCR-independent activation primarily induces the release of IFN-γ, Perforin, and Granzyme B, regulated by the transcription factors PLZF and T-bet, which are linked to a Tc1-like phenotype [30].
Studying these MAIT cell functional markers offers therapeutic potential in infectious diseases, cancer, and autoimmune disorders. Modulating TCR-dependent/independent, or cytokine-driven activation pathways could enhance immune responses or control inflammation, making MAIT cells promising candidates for immunotherapy across a range of conditions.
Transcription regulationUpon activation, MAIT cells exhibit a robust production of cytokines and activation markers. This dynamic response is tightly regulated by specific transcription factors, which trigger the downstream signaling pathways responsible for MAIT cell effector functions (Table 2). A key regulator of MAIT cell function is the transcription factor RORγt, which controls the production of IL-17A [34]. Beyond MAIT cells, RORγt is essential for the differentiation of other immune cells involved in type 17 immunity, such as Th17 cells, gamma delta T cells, and type 3 innate lymphoid cells, highlighting its broad role in regulating immune responses [35]. Although RORγt expression is nearly universal among MAIT cells, only a small fraction of these cells produces IL-17A ex vivo, suggesting either a pre-committed IL-17-producing subset or a broader potential for IL-17 production under the appropriate conditions [14, 36].
Table 2 Key transcription factors, functional significance, and associated signaling pathways of human MAIT cellsPLZF plays a pivotal role in shaping the innate-like features of MAIT cells. Expressed early during MAIT cell development, PLZF establishes an effector-memory phenotype in these cells. It acts as a master transcription factor, guiding MAIT cells toward their characteristic rapid-response behavior upon activation [37]. PLZF is consistently expressed across MAIT cell subsets, including CD8+, CD4−CD8− double negative (DN), and CD4+ cells, underscoring its central role in the functional maturation and maintenance of MAIT cells within the immune system [37].
T-bet is another critical transcription factor that modulates MAIT cell function, particularly shaping their Th1-like responses. During TCR-mediated activation, T-bet drives the transcription of genes involved in cytotoxicity and pro-inflammatory signaling, primarily promoting the production of IFN-γ, thereby strengthening MAIT cells’ ability to generate a robust immune defense. T-bet expression is notably higher in CD8+ MAIT cells compared to DN MAIT cells, highlighting its role in cytotoxic responses [38].
Eomesodermin, closely related to T-bet, is highly expressed in CD8+ MAIT cells and is pivotal for their cytotoxic function. It regulates the expression of cytotoxic molecules such as Granzyme B and Perforin, essential for MAIT cells’ ability to eliminate infected or malignant cells [39]. Upon activation, eomesodermin ensures the transcription of these effector molecules, thus linking TCR engagement to the cytotoxic arm of MAIT cell function. Unlike in peripheral blood, eomesodermin expression in mucosal MAIT cells, such as those in the endometrium, is more evenly distributed between CD8+ and DN subsets, suggesting a tissue-specific regulation of this transcription factor [38].
The transcriptional landscape of MAIT cells is not only dictated by their activation state but also by their tissue localization. Comparative analyses of MAIT cells in blood and tissue reveal distinct transcriptional profiles, particularly in organs such as the liver. Liver-resident MAIT cells demonstrate enhanced activity of the transcription factors AP-1 and NF-κB, both of which are instrumental in regulating inflammation and cytokine production [38]. AP-1, which includes proteins like FOS and JUN, and NF-κB are significantly upregulated in liver MAIT cells compared to their counterparts in the bloodstream. This elevation correlates with increased expression of genes associated with T cell activation, suggesting that liver-resident MAIT cells are primed for rapid immune responses. Additionally, the TCR-induced transcription factor EGR1 and RUNX3, which regulates tissue-resident memory cell differentiation, are more active in liver MAIT cells, further emphasizing the tissue-specific adaptations of these cells [40].
The transcriptional regulation of MAIT cell activation is a complex, multifaceted process governed by a combination of factors such as RORγt, PLZF, T-bet, and eomesodermin. These transcription factors not only define the cytokine and cytotoxic responses of MAIT cells but also facilitate their adaptation to specific tissue environments. Understanding how these transcriptional programs cooperate in different contexts provides valuable insights into the diverse roles of MAIT cells in immune surveillance, inflammation, and tissue-specific immunity.
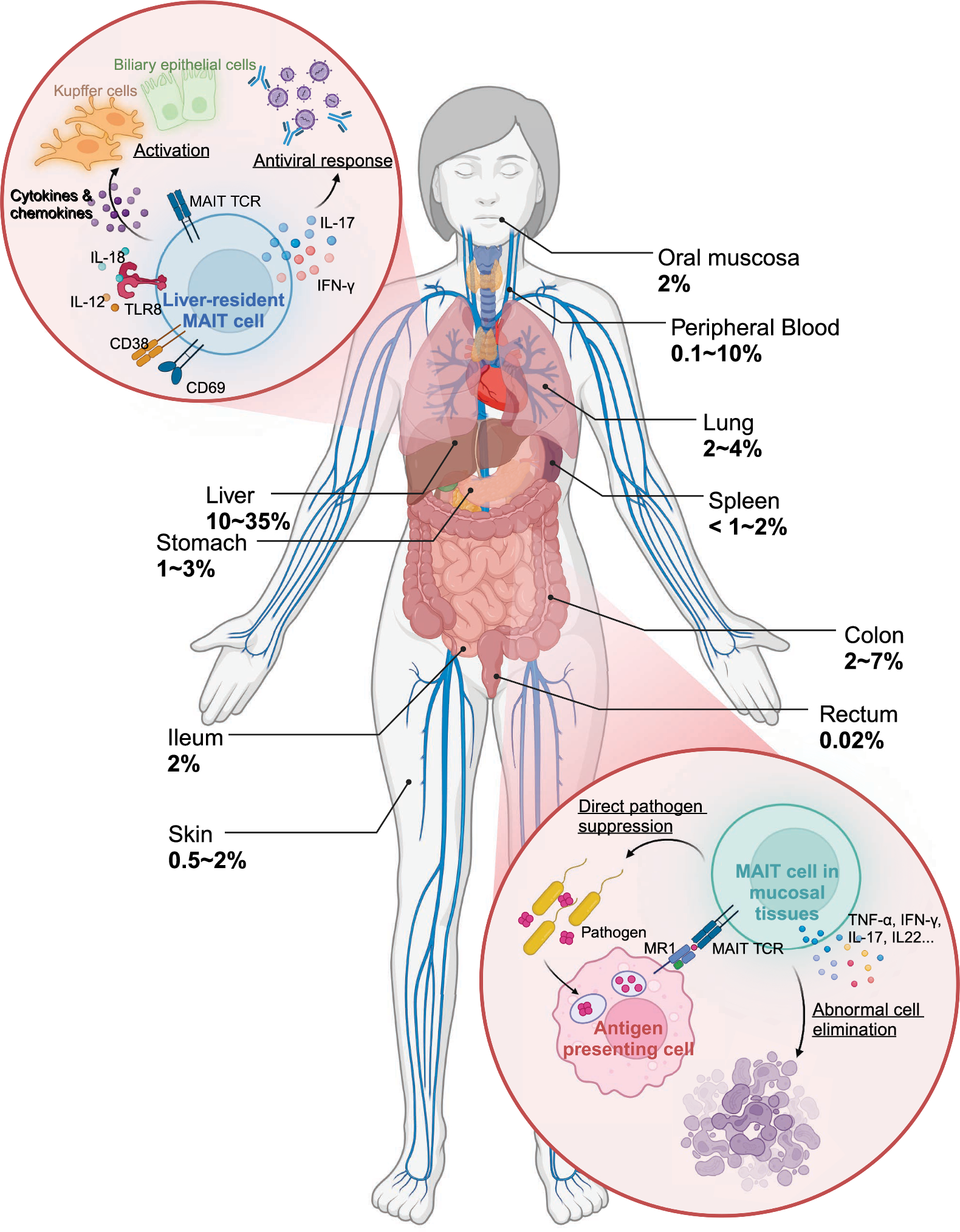
Tissue localization and immune surveillance of human MAIT cellsThe distinct localization of MAIT cells across barrier and mucosal tissues is crucial to their function in immune surveillance. These cells are particularly enriched in mucosal sites, such as the gut, lungs, and liver, where they account for 20–40% of T cells compared to their smaller presence (1–4%) in peripheral blood [11, 41]. Their high abundance at these sites underscores their specialization in detecting microbial threats and preserving tissue integrity.
Studies suggest that the tissue-specific distribution of MAIT cells is guided by the chemokine receptor CCR6, which binds to its ligand CCL20 expressed in mucosal tissues and the liver [42]. This receptor-ligand interaction ensures their recruitment to key sites of immune surveillance, where their functionality is tailored to the local environment. In the liver, MAIT cells exhibit a distinct pattern of distribution, being dispersed throughout the sinusoidal environment and portal fields, unlike other T cells, which are more localized to specific regions [42]. Liver-resident MAIT cells express activation markers, including CD69, HLA-DR, and CD38, which prepare them for rapid responses to microbial and inflammatory stimuli [43,44,45]. Functionally, they secrete IFN-γ and IL-17 in response to IL-12 and IL-18 stimulation during TLR8 activation, contributing to antiviral immunity and the regulation of hepatic inflammation and fibrosis [43, 45, 46]. As the predominant IL-17-producing T cell population in the liver, MAIT cells drive hepatic inflammation and fibrosis by activating Kupffer cells and biliary epithelial cells to release proinflammatory cytokines and chemokines [43, 46]. These functions underscore their critical role in liver immunosurveillance and regulation of immune responses to pathogens and tissue injury.
Beyond microbial defense, MAIT cells contribute to maintaining mucosal tissue integrity. In the gut, they produce IL-22 and IL-17, which are vital for epithelial repair and barrier maintenance [6]. IL-22 promotes the survival and proliferation of epithelial cells and stimulates mucus production by goblet cells, thereby reinforcing the physical barrier against pathogens [47, 48]. Meanwhile, IL-17 regulates tight junction proteins, such as occludin, to minimize barrier permeability during epithelial damage [49]. These cytokines collectively strengthen the mucosal barrier and limit excessive inflammation by suppressing the activation of conventional proinflammatory T cells, such as Th1 and Th17 subsets [50]. Similar functions have been observed in other mucosal tissues, including the genital tract, where MAIT cells play a role in epithelial repair and maintenance [24].
Collectively, MAIT cells are uniquely positioned in barrier and mucosal tissues, where they act as key mediators of immune surveillance and response (Fig. 1). Their ability to produce a diverse array of cytokines and cytotoxic mediators enables them to combat microbial threats, maintain epithelial integrity, and regulate local immune activity. However, there are still knowledge gaps regarding the specific mechanisms driving MAIT cell localization in tissues. One promising approach to address this is studying the role of tissue-specific cytokines in polarizing MAIT cells toward pro-inflammatory or regulatory responses. For instance, exposing MAIT cells from various tissues to different cytokine combinations (e.g., IL-12/IL-18, IL-15/IL-18, IL-10, TGF-β) can reveal how the balance of pro-inflammatory and regulatory cytokines influences their function and role in inflammation [51]. Additionally, exploring the dysregulation of MAIT cell activation and tissue homing in autoimmune disease pathogenesis offers another critical avenue. In multiple sclerosis, for example, MAIT cells infiltrate the brain, producing pro-inflammatory cytokines that may contribute to neuroinflammation [52]. In IBD, although MAIT cell frequency is reduced in peripheral blood, they accumulate in inflamed mucosa, suggesting a role in intestinal inflammation and tissue repair [53]. However, MAIT cells may also contribute to the pathogenesis of IBD. Upon activation, MAIT cells upregulate NKG2D, an activation receptor for NK cells, which further induces the expression of proinflammatory cytokines [8]. Clinical trials have shown that treatment with anti-NKG2D mAb can induce clinical remission in some patients with CD, highlighting the role of NKG2D in CD pathogenesis [54]. Additionally, in an oxazolone-induced mouse model of colitis, inhibiting MAIT cell activation through MR1 knockout or administration of the MR1 antagonist isobutyl 6-formylpterin has been shown to mitigate the severity of UC [55]. Studies focusing on the mechanisms of MAIT cell recruitment and their production of regulatory cytokines like IL-10 and IL-22 could provide valuable insights into their protective roles in autoimmune diseases.
Fig. 1
Tissue distribution and biological function of human MAIT cells. This figure illustrates the distribution of MAIT cells across various tissues in the human body. It shows the percentage of MAIT cells in each tissue, highlighting their localization and unique functional characteristics within the tissue microenvironment
Interactions between human MAIT cells and the microbiotaThe immune system and the microbiota maintain a close, mutually dependent relationship. As a crucial part of the immune system, MAIT cells mainly respond to bacteria through TCR recognition of metabolites generated in the vitamin B2 (riboflavin) biosynthetic pathway [5]. Studies have demonstrated that disrupting or inhibiting riboflavin metabolism in various bacteria suppresses MAIT cell responses, confirming that this pathway produces antigens targeted by MAIT cells [56,57,58,59]. This connection demonstrates the selective nature of MAIT cell responses and their reliance on specific bacterial metabolic pathways.
Early-life exposure to the microbiota is crucial in shaping MAIT cell development and function. Research in germ-free mice has demonstrated that MAIT cells are absent or underdeveloped without microbial exposure [60]. Additionally, the timing of microbial colonization plays a critical role in MAIT cell maturation. For instance, recolonizing neonatal germ-free mice with early-life intestinal commensal bacteria before the third week of life promotes MAIT cell development, whereas colonization beyond this critical window fails to trigger the same result [60, 61]. This time-sensitive interaction highlights a developmental window during which exposure to specific microbial signals is essential for effective MAIT cell function. Although the microbiota composition differs between humans and mice, the principles governing MAIT cell development appear conserved [62]. Both species require exposure to microbial-derived riboflavin metabolites for MAIT cell maturation. Variability in early-life microbial exposures, influenced by factors like delivery mode and environment, likely contributes to the diversity in MAIT cell abundance observed in humans. Insights from germ-free mouse models highlight the translational potential of targeting riboflavin metabolites to modulate MAIT cell function in infections and inflammatory conditions.
Beyond development, MAIT cells are essential for maintaining mucosal homeostasis and ensuring protection against infections. They are strategically positioned at barrier sites, such as mucosal surfaces, where they provide localized immune surveillance and promote tissue repair following microbial disturbances [61]. For example, MAIT cells promote tissue repair in response to microbial disturbances. Through MR1-mediated presentation of riboflavin, MAIT cells recognize skin commensals, facilitating cutaneous wound healing and contributing to homeostasis. In the presence of commensal organisms, these cells activate tissue-repair pathways, supporting the
Comments (0)