
Remember me
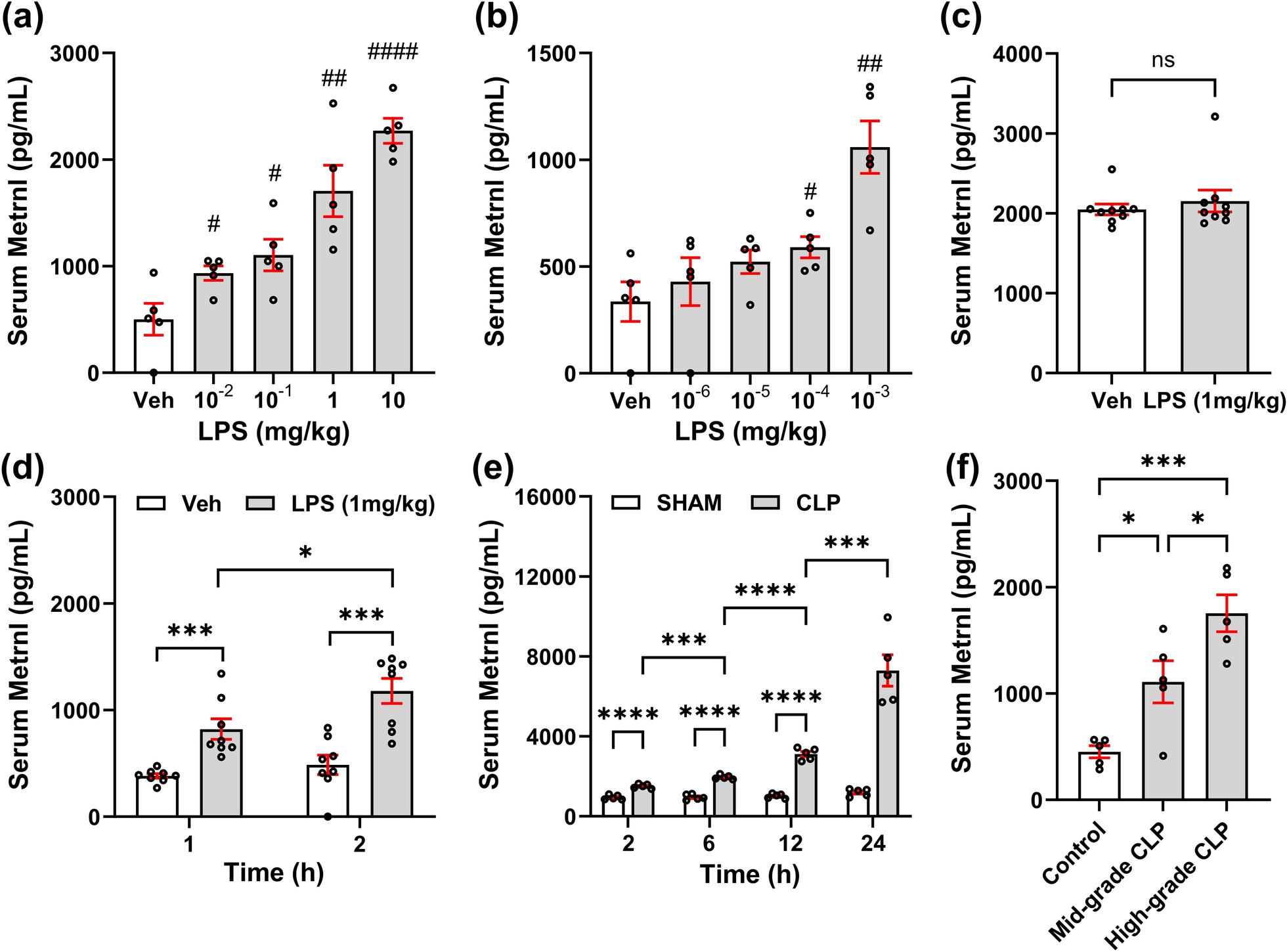





To investigate the relationship between sepsis and Metrnl levels, we utilized an LPS-induced sepsis model. Initially, different doses of LPS were administered to simulate different severities of sepsis. Two hours postmodeling, the experimental groups treated with LPS doses ranging from 10–2 to 10 mg/kg presented behavioral manifestations, including hypovolemia, fatigue, slow movement, and observable hemoconcentration during blood collection. As depicted in Fig. 1a, the serum levels of Metrnl increased in a dose-dependent manner with increasing LPS dosage. Additionally, there was a significant difference in the serum Metrnl levels between all the LPS groups and the Veh group. Further exploration with lower doses of LPS gradients revealed that doses as low as 10–4 mg/kg could still increase serum Metrnl levels (Fig. 1b).
Fig. 1
Metrnl is elevated during the early stage of sepsis in experimental mouse models. The mice were intraperitoneally injected with 10–2 to 10 mg/kg (a) or 10–6 to 10–3 mg/kg (b) of LPS or saline as a vehicle (Veh) control. Serum samples were collected 2 h later for Metrnl measurement. n = 5, #P < 0.05, ##P < 0.01, ####P < 0.0001 vs. Veh; (c, d) Mice were intraperitoneally injected with 1 mg/kg of LPS or saline as a vehicle control. Serum samples were collected 0.5 (c), 1 or 2 h (d) later for Metrnl measurement. n = 8, ns: no significance, *P < 0.05, ***P < 0.001; (e) Mice underwent CLP modeling, and serum samples were collected 2 to 24 h post-modeling for Metrnl measurement. n = 5, ***P < 0.001, ****P < 0.0001; (f) Mice underwent CLP modeling with different severity levels, and serum samples were collected 2 h post-modeling. The Control group was untreated. n = 5, *P < 0.05, ***P < 0.001
To investigate the diagnostic performance of Metrnl in sepsis, we examined the temporal relationship of serum Metrnl levels following the administration of a fixed dose of LPS (1 mg/kg). At 30 min postmodeling, there was no significant difference in the serum Metrnl levels between the LPS and Veh groups (Fig. 1c). However, as time progressed, a significant difference in the serum Metrnl levels between the LPS group and the Veh group emerged at 1 h postmodeling, and this difference further increased as time increased to 2 h (Fig. 1d).
We also employed a CLP-induced sepsis model, which better recapitulates the intricate progression of sepsis. Serum Metrnl levels were assessed at 2 to 24 h post-CLP modeling (Fig. 1e). At each time point, the serum Metrnl levels in the CLP group were consistently greater than those in the SHAM group. Moreover, with prolonged duration, the serum Metrnl levels in the CLP group tended to increase. Additionally, we compared serum Metrnl levels in mice subjected to different severity levels of CLP (Fig. 1f). Serum samples were collected 2 h post-CLP modeling. The serum Metrnl levels in the High-grade CLP group were significantly higher than those in the Mid-grade CLP group, suggesting a positive correlation between serum Metrnl levels and sepsis severity in the CLP model.
Serum METRNL has high diagnostic efficiency for sepsis in the ICUTo evaluate the potential of the METRNL as a biomarker, we compared the serum levels of METRNL, PCT, and CRP between patients with sepsis and non-sepsis controls. A total of 202 patients were recruited, including 107 sepsis patients and 95 non-sepsis patients. The clinical information of the participants is presented in Table 1. There were no significant statistical differences in sex or age distribution between sepsis patients and non-sepsis controls.
Table 1 Characteristics of patients with sepsis and controlsAs shown in Fig. 2a–c, the serum levels of METRNL, PCT, and CRP were increased in patients with sepsis in the ICU. Specifically, the mean value of METRNL increased from 443.18 ± 20.95 in control patients to 1031.16 ± 33.59 in sepsis patients, representing an increase of approximately 2.33 times. The diagnostic ability of biomarkers is typically evaluated via receiver operating characteristic (ROC) curves, where a larger area under the curve (AUC) indicates greater diagnostic accuracy. ROC curves for METRNL, PCT, and CRP analysis (Fig. 2d) revealed that METRNL and PCT had similarly high diagnostic accuracies for diagnosing sepsis, whereas CRP was less efficient (PCT: AUC 0.955, 95% CI 0.929–0.981; METRNL: AUC 0.943, 95% CI 0.91–0.975; CRP: AUC 0.873, 95% CI 0.825–0.921). Generally, the maximal Youden index determines the optimal diagnostic cutoff value [29]. In our study, the optimal cutoff values for METRNL, PCT, and CRP were determined to be 681.685 pg/mL, 0.5035 ng/mL, and 49.36 mg/L, respectively (Fig. 2e). At the optimal cutoff value, the sensitivities of METRNL, PCT, and CRP were all approximately 0.9. In terms of specificity, both the METRNL and PCT were also approximately 0.9, whereas the CRP was less than 0.7. Generally, METRNL and PCT demonstrated superior diagnostic performance, whereas CRP showed relatively poor diagnostic performance because of its lower AUC value and specificity.
Fig. 2
Serum METRNL has high diagnostic efficiency for sepsis in the ICU. Comparison of serum METRNL (a), PCT (b), and CRP (c) levels between sepsis patients (n = 107) and control patients (n = 95). **P<0.01, ***P<0.001; (d) AUC of serum METRNL, PCT, and CRP for sepsis diagnosis; (e) Diagnostic performance of METRNL, PCT and CRP for sepsis diagnosis. The percentage of sepsis patients detected positive by each biomarker was calculated to show the sensitivity, which was 0.897 (96/107) for METRNL, 0.925 (99/107) for PCT and 0.925 for CRP (99/107). The percentage of non-sepsis patients detected negative by each biomarker was calculated to show the specificity, which was 0.916 (87/95) for METRNL, 0.884 (84/95) for PCT and 0.695 (66/95) for CRP; The continuous changes of SOFA score (f), serum METRNL (g), PCT (h), and CRP (i) before and after sepsis in a single patient. Day 0 represents the time before sepsis onset, followed by data collection every 2 days
In addition, we were fortunate to collect continuous data from a sepsis patient spanning 30 days (Fig. 2f–i), with samples collected every two days, covering the period from pre-sepsis to post-sepsis onset. These longitudinal data provide a better understanding of the changes in various biomarkers during the progression of sepsis. The patient did not have sepsis on day 0. Two days later, the SOFA score indicated that the patient had developed sepsis, and at that time, the patient's serum METRNL, PCT, and CRP levels were approximately 4.5, 11, and 2.3 times the pre-sepsis levels, respectively. Additionally, the METRNL and PCT levels remained consistently elevated throughout the progression of sepsis, whereas the CRP level temporarily decreased below the baseline.
Serum Metrnl increases earlier than PCT and CRP in the LPS-induced sepsis modelTo further validate the potential of Metrnl as a biomarker for sepsis, we conducted a comparative analysis between Metrnl and established sepsis biomarkers in animal models. Two hours after LPS-induced sepsis modeling, the results revealed a significant increases in the serum Metrnl and PCT levels compared with those in the control group, whereas the serum CRP levels remained unchanged (Fig. 3a–c). Subsequently, investigations were also carried out at the 1 h post-modeling time point. Compared with those in the control group, the serum PCT and CRP levels were not significant different, whereas the Metrnl levels continued to exhibit a significantly increase (Fig. 3d–f). These findings indicate that, compared with classical markers such as PCT and CRP, Metrnl has greater sensitivity and responsiveness in the early stages of sepsis.
Fig. 3
Serum Metrnl rises earlier than PCT and CRP in LPS-induced sepsis model. The mice were intraperitoneally injected with 1 mg/kg of LPS or saline as a control and serum samples were collected 2 h (a–c) or 1 h (d–f) after injection. n = 5, ns: no significance, **P < 0.01
Serum METRNL is derived mainly from endothelial cells under septic conditionsTo determine whether serum METRNL is secreted primarily by endothelial cells under septic conditions, mice with endothelial cell-specific knockout of Metrnl (EC-Metrnl−/−) were generated by mating MetrnlloxP/loxP mice with Tek-Cre mice (Fig. 4a, b). As shown in Fig. 4c, the serum Metrnl levels in EC-Metrnl−/− mice in the Veh group were significantly lower than those in WT mice, and the serum Metrnl level was below the detection limit, suggesting that the serum Metrnl secretion is derived mainly from the vascular endothelium under physiological conditions. Next, we investigated the primary source of serum Metrnl during sepsis in both the LPS-induced and CLP-induced sepsis mouse models. In the LPS-induced sepsis mouse model, the serum Metrnl level in the experimental group of EC-Metrnl−/− mice was approximately 18% of that in WT mice (Fig. 4c). We subsequently established CLP sepsis models in EC-Metrnl−/− and WT mice and blood samples were collected 24 h after modeling to detect serum Metrnl. All the mice exhibited hypovolemia and hemoconcentration. As shown in Fig. 4d, the serum Metrnl level in sham EC-Metrnl−/− mice was approximately 10% of that in WT mice, and the serum Metrnl level in CLP-modeled EC-Metrnl−/− mice was approximately 6% of that in CLP-modeled WT mice. Additionally, compared with those in the corresponding control groups, the mean serum Metrnl levels in WT mice increased by 757.94 pg/mL after LPS modeling and by 5273.27 pg/mL after CLP modeling. In contrast, the increases in those of EC-Metrnl−/− mice were only 254.71 pg/mL and 288.26 pg/mL, respectively, indicating that the elevation of serum Metrnl levels in the sepsis model mice was significantly reduced after endothelial cell-specific knockout of Metrnl. These results indicate that the vascular endothelium is the primary cell source of serum Metrnl in the septic state.
Fig. 4
Serum METRNL is derived mainly from endothelial cells under septic conditions. (a) Endothelial cell (EC)-specific Metrnl knockout (EC-Metrnl−/−) mice were generated by mating MetrnlloxP/loxP and Tek-Cre mice; (b) Identification of the EC-Metrnl−/− mice. 1, 2, 5: MetrnlloxP/loxP Tek-Cre (EC-Metrnl−/−); 3, 4, 6: MetrnlloxP/loxP (WT); The PCR products of MetrnlloxP, Tek-Cre, and the internal reference gene are 243 bp, 324 bp, and 100 bp, respectively; (c) EC-Metrnl−/− mice and WT mice were intraperitoneally injected with 1 mg/kg LPS or saline as Veh control. Serum samples were collected 2 h after injection; (d) EC-Metrnl−/− mice and WT mice underwent CLP modeling or sham surgery. Serum samples were collected 2 h after modeling. n = 5, **P < 0.01, ***P < 0.001, ****P < 0.0001; (e) HUVECs were stimulated with 10–3 to 10 mg/mL LPS or Veh control for 2 h, and the cell supernatants were collected for METRNL measurement. n = 6, ####P < 0.0001 vs. Veh; (f) HUVECs were stimulated with 1 mg/mL LPS or vehicle control for 0.5 to 24 h, and cell supernatants were collected for METRNL measurement. n = 6, **P < 0.01, ****P < 0.0001
Furthermore, we explored the dose–response and time-response relationships of METRNL levels in an LPS-induced sepsis cell model. Initially, HUVECs were treated with different concentrations of LPS, and METRNL levels in the cell supernatant were uniformly measured 2 h post-treatment. As depicted in Fig. 4e, compared with the Veh group, the group treated with LPS at concentration as low as 10–3 μg/mL presented in nearly doubled METRNL levels in the cell supernatant. Furthermore, with increasing LPS incubation concentrations (from 10–3 to 10 μg/mL), the METRNL level in the cell supernatant increased in a dose-dependent manner. The cells were subsequently incubated with an equal concentration of LPS (1 μg/mL), and the METRNL levels in the cell supernatant were measured at different time points ranging from 0.5 to 24 h post-treatment. Treatment with LPS for 30 min induced a significant increase in METRNL in the supernatant. The METRNL levels in the LPS groups were significantly different from those in the corresponding Veh groups. A time-response relationship between METRNL levels and the duration of LPS exposure was also observed (Fig. 4f).
METRNL is secreted via the classical ER-Golgi pathway under LPS stimulationIn the classical endoplasmic reticulum-Golgi (ER-Golgi) pathway, proteins are transported from the endoplasmic reticulum to the Golgi apparatus via COP II vesicles, and all three components are essential [30,31,32]. First, we used two inhibitors (Brefeldin A and Eeyarestatin 1) to validate the involvement of the endoplasmic reticulum and Golgi apparatus in the secretion of METRNL under LPS stimulation. Brefeldin A blocks the transfer of newly synthesized proteins from the endoplasmic reticulum to the Golgi apparatus [33]; Eeyarestatin 1 inhibits the construction of ER membrane transport channels [34]. In the Brefeldin A group, the METRNL levels in the HUVEC supernatant were approximately 10% of those in the solvent control group, whereas in the Eeyarestatin 1 group, the METRNL levels were approximately 20% of those in the solvent control group (Fig. 5a). Additionally, upon removal of Brefeldin A, the release of METRNL from endothelial cells was fully restored (Fig. 5b). We further applied these two inhibitors under septic conditions. The cells were treated with LPS and Eeyarestatin 1 or Brefeldin A for 2 h. As shown in Fig. 5c, the level of METRNL in the supernatant significantly increased in the Veh + LPS group. Moreover, treatment with Eeyarestatin 1 or Brefeldin A both blocked the LPS-induced increase in METRNL levels in the supernatant. These results indicated that the endoplasmic reticulum and Golgi apparatus were involved in the secretion of METRNL under LPS stimulation.
Fig. 5
METRNL is secreted via the classical ER-Golgi pathway under LPS stimulation. (a) METRNL secretion from primary cultured HUVECs was reduced by Eeyarestatin 1 (5 mg/mL) or Brefeldin A (5 mg/mL). n = 6; (b) Recovery of METRNL release from HUVECs after Brefeldin A removal. Brefeldin A (5 mg/mL) or a vehicle control (DMSO) was added to the serum-free culture medium 2 h before cell supernatant collection for METRNL measurement. Then the cells were washed for three times and incubated in normal culture medium without drugs for 16 h followed by another 2 h of incubation in serum-free medium before METRNL detection. n = 6; (c) METRNL secretion from HUVECs induced by LPS (1 mg/mL) stimulation was inhibited by Eeyarestatin 1 (5 mg/mL) or Brefeldin A (5 mg/mL). n = 4; (d, e) After transfection with SAR1A/B siRNA, the mRNA expression of SAR1A (d) and SAR1B (e) was reduced in HUVECs; (f) METRNL secretion from HUVECs induced by LPS (1 mg/mL) stimulation was reduced by transfection with SAR1A/B siRNA. n = 4. ns: no significance, **P < 0.01, ***P < 0.001, ****P < 0.0001
Second, we investigated the involvement of COP II vesicles. The small G protein SAR1, which contains two isoforms, SAR1A and SAR1B, is a critical component in the formation of COP II vesicles [35]. Therefore, we transfected HUVECs with SAR1A/B siRNA, which targets the two different isoforms of SAR1, to validate whether COP II vesicles are involved in METRNL secretion under septic conditions. RT-PCR revealed that the SAR1A knockdown efficiency was approximately 84.18% (Fig. 5d), and the SAR1B knockdown efficacy was approximately 89.88% (Fig. 5e). Transfection of SAR1A/B siRNA in HUVECs blocked the increase in METRNL levels in the supernatant induced by LPS and resulted in a decrease in the secretion levels of METRNL under normal conditions. (Fig. 5f). These results collectively demonstrated that METRNL was primarily secreted via the classical ER-Golgi pathway under LPS stimulation.
LPS stimulation promotes the secretion of METRNL through TLR4 activation of downstream ERK kinase.Toll-like receptor 4 is known for recognizing LPS and promoting inflammation by initiating the release of cytokines and chemokines [36,37,38]. To investigate the mechanism of LPS-induced METRNL release, we utilized TAK-242 [39, 40], a Toll-like receptor 4 (TLR4) inhibitor, to treat both animals and cells. In mice, TAK-242 (3 mg/kg) was intraperitoneally injected 2 h before LPS modeling, and blood samples were collected from the heart 2 h after modeling to measure serum Metrnl levels. As shown in Fig. 6a, the increase in mouse serum Metrnl induced by the intraperitoneal injection of LPS was reduced by TAK-242. Consistent with the results from the animal experiments, pretreatment of HUVECs with TAK-242 (1 μM) for 2 h followed by LPS administration and further incubation for 2 h resulted in significantly lower levels of METRNL in the cell supernatant in the TAK-242 + LPS group than in the DMSO + LPS group, indicating that TAK-242 blocked the LPS-induced increase in METRNL in the cell supernatant (Fig. 6b).
Fig. 6
LPS stimulation promotes the secretion of METRNL through TLR4 activation of downstream ERK kinase. (a) TAK-242 reduced serum Metrnl increase induced by LPS modeling. Mice were pretreated with an intraperitoneal injection of TAK-242 (3 mg/kg) for 2 h, followed by an intraperitoneal injection of LPS (1 mg/kg). Serum were collected 2 h after injection. n = 5; (b) The increase in METRNL secretion from HUVECs induced by LPS (1 mg/mL) stimulation was inhibited by pretreatment with TAK-242 (1 μM) for 2 h. n = 4; The increase in METRNL secretion (c) and phosphorylation (d) from HUVECs induced by LPS stimulation (1 mg/mL) was inhibited by pretreatment with U0126 (10 μM) for 2 h. n = 4; (e) The levels of ERK phosphorylation in HUVECs increased with higher concentrations of LPS. ERK phosphorylation levels were measured 30 min after stimulation with LPS. n = 6; (f) Secretion and regulatory mechanism of METRNL in sepsis condition (By Figdraw). ns: no significance, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001
As an important group of signal transduction proteins, the ERK family regulates crucial processes such as cell growth, development, division, and death [41]. It has been reported that ERK is associated with protein secretion [42]. Therefore, we used the ERK phosphorylation inhibitor U0126 to investigate whether ERK acts as a key node protein connecting the TLR4 pathway and the ER-Golgi secretion pathway. The results revealed that using U0126 (10 μM) significantly reduced the increase in METRNL secretion induced by LPS stimulation (Fig. 6c), whereas a decrease in ERK phosphorylation was observed (Fig. 6d). Additionally, the phosphorylation of ERK increased with increasing doses of LPS (Fig. 6e). Overall, we concluded that the LPS-stimulated increase in METRNL secretion was mediated via the TLR4-ERK pathway.
Comments (0)