Autism spectrum disorders (ASD) are a group of life-long neurodevelopmental disorders marked by difficulties in social interactions, along with restricted, repetitive behaviours and interests. Symptoms typically emerge in early childhood and can vary in severity. They often intensify during development, resulting in clinically significant impairments in daily functioning [1].
Interestingly, the prevalence of ASD has increased significantly across various countries over the past few decades [2, 3] placing a substantial health and economic burden on communities worldwide. Although factors like advanced maternal and paternal age or maternal metabolic conditions during pregnancy may have contributed to the rise in ASD prevalence [4,5,6], heightened awareness and improved diagnostic capabilities are likely the predominant drivers of the current trend [7].
However, despite increased awareness and progress in diagnostics, the development of adequate treatment methods lags behind. Insufficient funding for research and services as well as a lack of knowledge on evidence-based treatments have been identified as significant gaps in ASD care [8]. This is unsurprising, as evidence-based treatments for ASD are scarce. Among the multitude of treatment modalities, only a few behavioural interventions and medical therapies have been proven effective [9, 10], primarily in children up to 12 years of age as other age groups remain underrepresented in research [11]. Yet, behavioral interventions are often time- and cost-intensive [12], while psychopharmacological treatments largely target comorbid conditions [13] and carry an increased risk of side effects for children and adolescents with ASD [14]. It is therefore crucial to allocate more resources toward the exploration of treatment alternatives, while also expanding the study population beyond children.
As the symptomatology of ASD varies significantly across affected individuals [15], research on treatment methods proves somewhat difficult. Thus, focusing on underlying mechanisms, particularly neurophysiological alterations, rather than solely targeting behavioral manifestations, may offer a more effective framework for understanding and addressing the disorder [16].
Consistent evidence from structural magnetic resonance imaging (MRI) studies in ASD (summarized in Patil et al. [17]), reports a reduction in gray matter volume in regions crucial for social interactions and emotional processing, including the amygdala, the fusiform gyrus and the superior temporal sulcus. Regarding functional connectivity, research has shown reduced connectivity within the default mode network, a brain network implicated in self-reflection and theory of mind. These selected findings demonstrate that brain areas involved in social, emotional, and sensory processing - functions often impaired in ASD - are indeed frequently found to be both structurally and/or functionally altered, indicating a neurobiological foundation of the disorder’s symptomatology.
While insights from MRI studies provide valuable information, electroencephalography (EEG) has a much higher temporal resolution and measures neural activity directly, highlighting its importance as a complementary method for understanding neural processes. Furthermore, EEG does not present specific challenges inherent to MRI, such as confined spaces, loud noises, and the need to remain still, that are frequently perceived as barriers by individuals with ASD [18]. In addition to being comparably inexpensive and broadly accessible, these advantages make EEG an essential tool for examining neural processes in ASD.
Resting-state (rs) EEG, which reflects an individual’s typical neural activity during rest (i.e., when not engaged in mentally or physically demanding tasks), is one of the most established EEG-based measures, alongside event-related potentials (ERPs). However, rsEEG recordings are conducted under conditions that are oftentimes heterogeneous, thereby complicating comparability across studies. rs paradigm types (eyes-closed (EC) vs. eyes-open (EO)) vary across studies, with differences in recording durations of EC and EO segments or the inclusion of only one condition, all of which can affect the recorded data [19]. Additionally, factors such as the day of the experiment, the time of day, and the level of physical activity prior to recording - variables likely differing across studies - have been shown to impact rsEEG data [20]. Moreover, significant heterogeneity in methodological approaches and the targeting of different EEG features across studies further complicates the comparison of results. However, despite these challenging conditions, certain abnormal power band characteristics have been repeatedly observed in individuals with ASD during rs measurements.
Regarding the Delta band, increased power is frequently reported, both in absolute [21,22,23,24] and relative measures [21, 22, 25]. This increased Delta power has been observed primarily in frontal areas [23, 24], but also throughout the entire cortex [22, 25].
Similarly, concerning Theta power, higher absolute [22, 23, 26] and relative [22, 27, 28] power has recurrently been assessed, particularly in frontal, prefrontal, and midline regions [22, 23, 26, 28].
Regarding the high-frequency Beta band, some studies suggest increases in both absolute [29, 30] and relative power [30]. Furthermore, some evidence suggests increased absolute [29,30,31] and relative Gamma power [30, 32], specifically in frontal, central and parietal regions [31, 32].
While multiple studies have shown increased power in the low-frequency (Delta and Theta) and high-frequency (Beta and Gamma) bands, the Alpha band exhibits a contrasting pattern. Reduced absolute and relative Alpha power is a consistent and robust finding across numerous rsEEG studies in ASD [21,22,23,, 22, 25, 28, 30, 33, 34, 35, 36].
Taken together, these findings suggest a U-shaped profile of rs power abnormalities in ASD, as outlined in a review by Wang et al. [37]. This profile is characterized by heightened power in both low and high frequency bands, specifically increased Delta and Theta power at the lower end, as well as Beta and Gamma power at the higher end and reduced Alpha activity, which is located in the lower-middle range of the power spectrum [37]. Despite some results contradicting the U-shaped curve hypothesis [24, 27, 38], likely due to ASD’s neurophysiological diversity and heterogeneous measurement conditions, certain power abnormalities remain consistent and robust when reviewing the current research. A recent systematic review and meta-analysis by Neo et al. [19], which included 41 studies with 1246 autistic and 1455 neurotypical individuals (with some overlap with Wang et al. [37]), partially supports the U-shaped curve hypothesis, identifying a reduction in relative rs Alpha power and an increase in both absolute and relative Gamma power in individuals with ASD. In addition, Neo et al. [19] demonstrated that the type of rs paradigm (EC vs. EO) significantly moderated effect sizes, with the EO condition producing larger effect sizes. Similarly, the duration of rs recordings moderated Alpha power effect sizes, with longer recording durations yielding more pronounced effects. These findings further suggest that the potential of rs Alpha power as a robust biomarker for ASD is particularly evident when measured under EO conditions with sufficiently long recording durations.
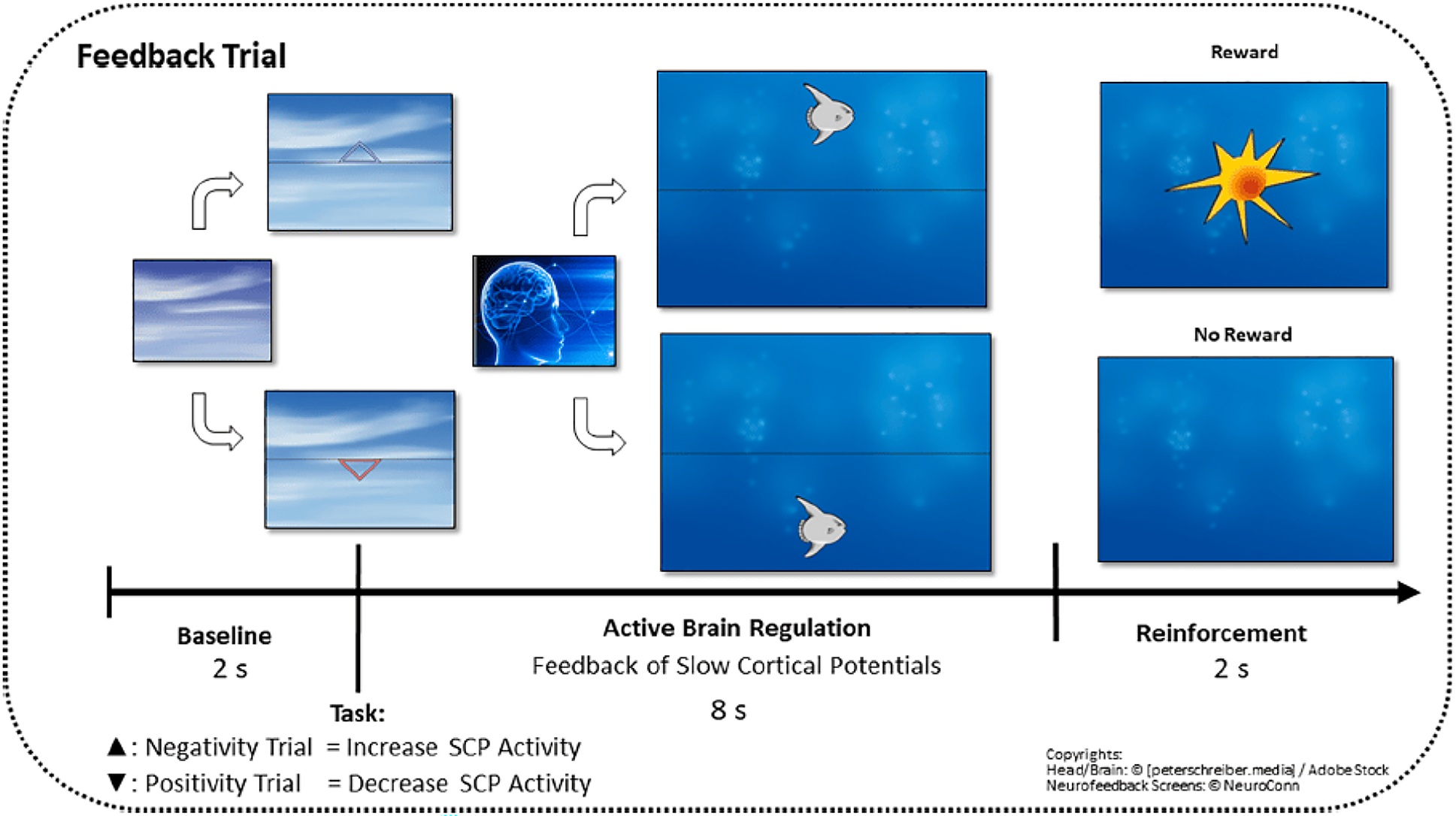
Although several electrocortical abnormalities have been identified in ASD, biological treatment approaches targeting neural mechanisms are still in their infancy. EEG neurofeedback (NF) is a safe, side-effect-free, biologically based treatment approach, ideally suited to address abnormal EEG activity. Research focused on children and adolescents has demonstrated its potential in conditions where such abnormalities are prevalent, including e.g. epilepsy [39, 40], anxiety [41], and trauma [42]. In the context of ASD, several recent reviews [43,44,45,46] have concluded that NF holds promise as a tool for alleviating ASD symptoms. However, as noted by Kumari & Sharma [43], the evaluation of these symptoms has predominantly relied on subjective instruments along with parental input. Given NF’s susceptibility to non-specific effects [46], incorporating objective measures alongside subjective assessments is necessary to validate its efficacy. This became particularly evident in attention deficit hyperactivity disorder (ADHD), where two recent meta-analyses found improvements in attentional performance following NF [47, 48]. However, Chung et al. [47] found no benefit over placebo when only subjective measures were used in double-blind studies, while Chiu et al. [48] observed significant improvements with objective behavioral measures. This underscores the importance of adding objective outcomes to address the limitations of subjective evaluations and provides further support for NF’s efficacy. As NF is grounded in biological principles, it is crucial to assess biologically based outcomes, extending beyond behavioral measures, to ensure a direct assessment of the underlying neural mechanisms affected by the treatment.
Indeed, studies have shown that NF can alter ERPs in both children [49] and adults with ADHD [50]. In ASD, Konicar et al. [51] demonstrated that adolescents were able to modify their brain activity through NF training. EEG recordings during training showed a reduction in Delta power and an increase in Alpha power. Moreover, these neurophysiological changes were associated with improvements in ASD symptoms, as measured by the Social Responsiveness Scale (SRS), further validating the potential of NF. However, it remains unassessed whether the applied NF- training also affected rsEEG activity as a neural signature frequently aberrant in ASD.
Changes in rsEEG activity after NF training have been assessed in various populations, such as learning-disabled children [52], adults with alcohol use disorder [53] and children with ASD [54], suggesting that NF can induce a reorganization of neural activity at rest, beyond localized adjustments at trained electrode sites. In ASD, Pineda et al. [54] found that children who underwent mu rhythm- NF training, showed decreased rs mu amplitude and coherence at central sites. Similarly, Kouijzer et al. [55] reported that children with ASD exhibited reduced rs Theta and Delta activity, along with increased resting Beta activity at central sites after having been trained with a Theta/Beta protocol. In a follow-up study, efforts to reduce Theta power in children with ASD were successful, showing decreased Theta power both during the sessions and in the post-treatment rsEEG measurements [56]. Notably, the decreases in Theta power during session data and in the rsEEG were highly correlated, suggesting that the reduction in Theta power induced by NF was maintained after treatment concluded.
Summarized, a key observation in the rsEEG literature in ASD is the consistent reduction in relative (particularly in EO paradigms [19]), as well as in absolute Alpha power [34,35,36] and an increased absolute Delta power [21,








Comments (0)