The results reported in this paper show that the majority of modern Polish Y-chromosomal lineages belong to young yet relatively large subclades that are common in other Slavic-speaking populations. This strongly supports the hypothesis of Early Slavic mass migrations dated to the middle of the first millennium CE and is consistent with previous studies demonstrating that modern populations of Central, Eastern and South-Eastern Europe share a very significant amount of recent genetic ancestry (Ralph and Coop 2013) that likely comes from an ancient population resembling modern Poles or Lithuanians (Hellenthal et al. 2014; Busby et al. 2015). It is also worth mentioning that a good correlation between the modern Y-chromosomal and autosomal data in this part of Europe has previously been noted by others (Kushniarevich et al. 2015), so the major conclusion from all these results is that Early Slavic migrations were not just a movement of relatively small male-dominated elites. Instead, the migration process likely involved much larger groupings with both male and female individuals equally contributing to the final outcome, as recently suggested based on ancient DNA data (Olalde et al. 2023).
In light of the above, it is important to ask why many mtDNA studies have failed to come to similar conclusions regarding the Early Slavic migrations. One of the likely reasons is that the analysis of mitochondrial DNA does not ensure sufficient resolution to easily discriminate between lineages diverging in relatively recent times, for example during the last 2–3 thousand years, thus in a period that seems to have been most crucial for the process of Slavic ethnogenesis. Since the coalescence age estimates for most mtDNA haplogroups investigated in this context significantly exceed 3 kya, it is extremely difficult to make conclusions about their local or non-local origin, especially when relatively recent migrations from not-so-distant European subregions need to be taken into consideration. Yet another obstacle is that the frequencies of all major mtDNA haplogroups are very similar in most of Europe, so any migrations between neighboring European regions are hard to follow using mtDNA data. To summarize, any conclusions suggesting genetic continuity since the Bronze or Iron Age that are based either on the presence of relatively old and widely spread haplogroups or on similar proportions of macro-haplogroups should be considered unjustified. However, despite all those limitations associated with using mtDNA data to trace migrations of Early Slavs, it is worth noting that many mtDNA subhaplogroups that have previously been suggested to be associated with the Slavs (like, for example, H5a2, H5e1a, U4a2 or U5a2b1 (Mielnik-Sikorska et al. 2013) have turned out to be either absent or very rare in the Polish Iron Age population, while being relatively frequent among Medieval Poles (Stolarek et al. 2023), which is consistent with the hypothetical Early Medieval influx of Slavic-associated mtDNA haplogroups from outside. At the same time, there is an interesting example of haplogroup U3a1a that was exceptionally frequent in the Iron Age Wielbark culture but absent among Medieval Poles and very rare in Modern Poland, suggesting a significant population replacement between Late Roman and Medieval times.
Although our results strongly favor the Early Slavic migration scenario, we cannot rule out that there were also other migrations that could have been responsible for the current distribution of at least some of those Y-chromosomal subclades that today are associated mostly with the Slavs. Certainly, the relatively broad confidence intervals of TMRCA estimates used in our study add some uncertainty regarding the exact time frames of suggested migrations. On the other hand, there were not many known large-scale migration processes in this part of Europe that could have been dated to the first millennium CE while covering about the same area as the migrations of Early Slavs. According to both archaeology and written records, the only known migration of similar extent could be be that of the ancient Gothic people who expanded from today’s Poland to the North Pontic region and then towards the Balkans during the Late Roman period. However, in such case one should expect that the migrations were associated mostly with haplogroup I1 that was extremely common among the Gothic people of the Wielbark culture in Poland (Stolarek et al. 2023), rather than with many different subclades of haplogroup R1a that have been identified in our study.
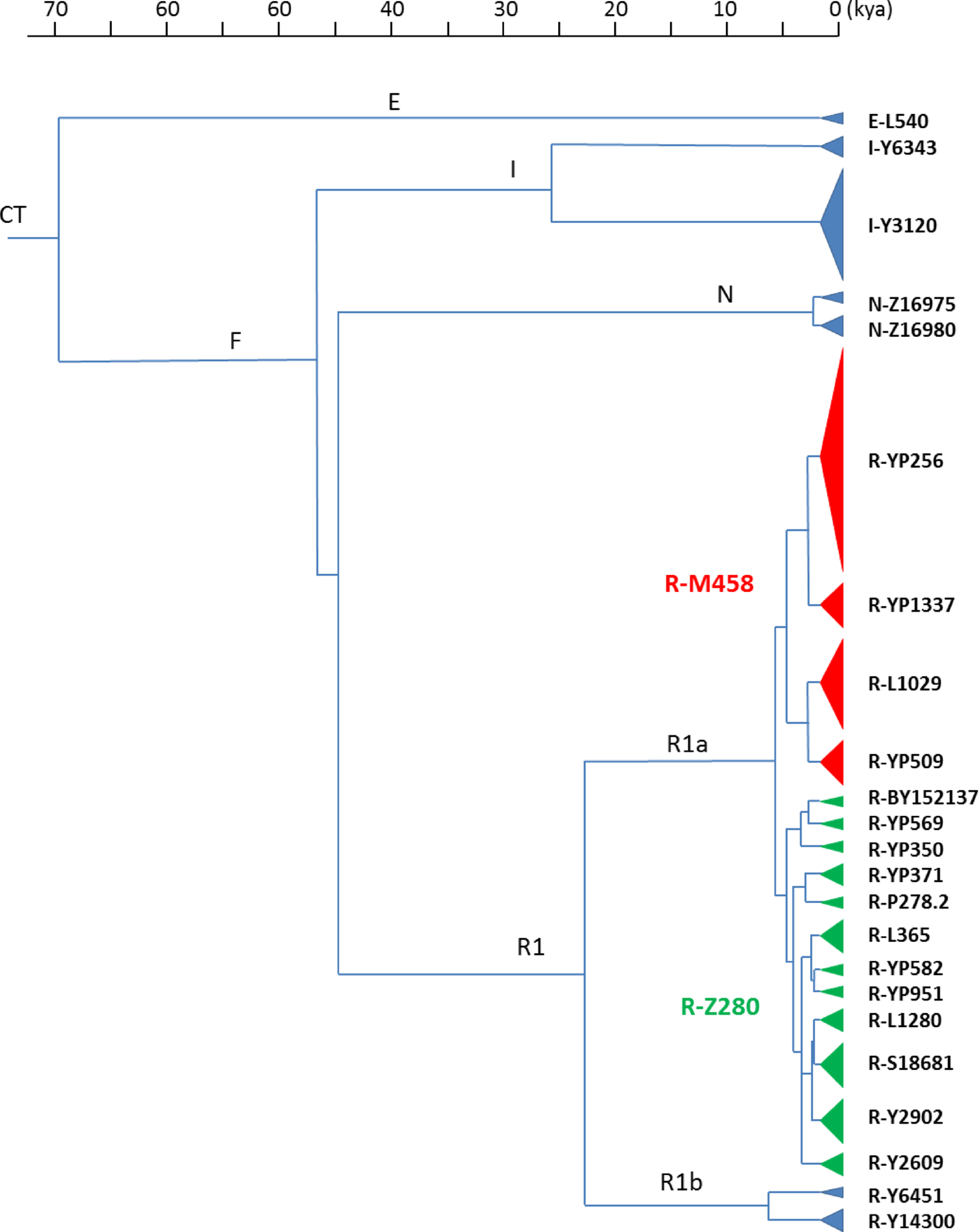
The Y-chromosomal haplogroup R1a and especially its Central-Eastern European subbranch R-M458 have long been suspected of being associated with the ethnogenesis of the Slavs (Underhill et al. 2010; Kushniarevich et al. 2012). Indeed, our results strongly support the often expressed view that haplogroup R1a was common among the Early Slavs. Furthermore, we demonstrate that two subhaplogroups R-M458 and R-Z280, together representing 96.9% (310/320) of all R1a samples in our group, constituted the core of the Early Slavic population. Intriguingly, although clade R-M458 included more samples in our study than clade R-Z280 (174 or 29.1% of total vs. 136 or 22.7%, respectively), all those R-M458 males could be assigned to just four large Iron Age-derived subclades. In contrast, the number of separate Iron Age-derived subclades of R-Z280 was much larger (12), yet these subclades were generally smaller and not all of them seem to be associated with deep Slavic ancestry, not to mention that a significant fraction of all R-Z280 members (19/136, 14.0%) have not been included in any of those 12 major subclades. This indicates that the closely related clades R-M458 and R-Z280 had substantially different demographic histories, hence future efforts should be focused on explaining whether those differences are related to different roles those two clades played in the formation of the hypothetical Late Proto-Slavic community.
Once we know which Y-chromosomal subclades were likely common among expanding Early Slavs, we should also be able to determine the location of the hypothetical Proto-Slavic homeland based on ancient DNA data. However, none of the Iron Age populations studied so far fit the picture suggested by the results presented in this paper. For example, none of the so far investigated Iron Age cultures in Central or Eastern Europe show elevated frequencies of both R-M458 and R-Z280. Also, although I-Y3120 seems to be the largest of all Slavic-associated subclades of Early Iron Age origin, not a single example of this subclade has been found in Pre-Medieval Europe, so far. All this suggests that the ancient Proto-Slavic population we are looking for has not been sampled yet. At this point, it is worth mentioning that although the exact location of the hypothetical homeland of the Early Slavs remains a subject of ongoing discussion, many archaeologists point to the Kiev culture (3-5th cent. CE) in Northern Ukraine and in the neighboring regions of Southern Belarus and Western Russia as the most likely source of the Early Slavs (Parczewski 2005; Kobyliński 2005; Kazanski 2020). Unfortunately, the dominant funerary practice in the Kiev culture, as well as in the preceding Zarubintsy culture, was cremation, which is one of the main reasons why ancient DNA results for the Iron Age populations residing in this particular part of Eastern Europe are still missing.
In modern Poland, the combined frequency of all 18 Slavic-associated Y-chromosomal subclades identified in our study is 58.4%, or 59.5% when including the seven members of the West Slavic-associated N-Y6075 subclade of Baltic clade N-Z16980. This number would likely increase when taking into account all potentially Slavic subclades (mostly those under R-Z280) that are relatively rare in Poland, and thus were not represented by at least three males in our study group, hence more detailed studies on different Slavic populations are needed to get a more precise estimate of the Early Slavic contribution to the modern Polish Y-DNA pool. The same is true for lineages descending from the local Iron Age population, as the combined frequency of subclades I-Y6343 and R-Z17913 (2.0%) likely represents only a part of the local pre-Slavic (Germanic) contribution. Nonetheless, the above numbers allow us to roughly estimate the ratio of Early Slavic newcomers to locals as between 60:40 and 97:3. Such a strong domination of immigrants over local residents is consistent with archaeological findings and historical reports suggesting significant depopulation of many regions in Poland during the Great Migration period (Pędziszewska et al. 2020; Wołoszyn 2020).
Apart from the Y-DNA lineages that have been brought to Poland with the expanding Early Slavs and those descending from the local Pre-Slavic population, our study group likely includes many lineages that have become part of the Polish Y-chromosomal pool in more recent times, i.e. mostly in the second millennium CE. This may, for example, include lineages brought to Poland with ethnic Germans and other people of Western European ancestry (including, for example, Dutch, Flemish or Scottish people) who have been settling in Poland since the Medieval times. Unfortunately, discriminating between lineages of deep Germanic or Celto-Germanic ancestry that were brought to Poland in Pre-Slavic times and those arriving significantly later is possible only in a situation where we know about either predominantly Polish or predominantly foreign (North-Western European) brother lineages diverging around the middle of the first millennium CE. In practice, this means that for rare lineages that are not part of any young yet relatively large subclade, such discrimination will be possible only after more people from relevant populations are tested.
Another significant group of foreign (non-Slavic) Y-chromosomal lineages that could have joined the Polish Y-DNA pool following the arrival of Early Slavs are subclades associated with deep Baltic ancestry. There seem to be two major groups of such Baltic-associated subclades, with one encompassing different subclades of N-L1025, a large Baltic-specific subhaplogroup that includes, among others, N-Z16975 and N-Z16980 (both well represented in our group of Polish males), and the other being composed of some relatively rare non-Slavic subclades of R-Z280, like R-YP350. Although the identification of all Baltic-associated subclades of R-Z280 requires much more detailed studies, it seems that the overall contribution of Baltic Y-chromosomal lineages in Poland is about 4–8%, with as much as about 3% corresponding to clade N-L1025 alone. However, it should be pointed out that this estimation does not take into account the possibility that some Early Slavic-associated subclades, especially those from haplogroup R1a, could have originally been associated with the neighboring Baltic-speaking people who may have been assimilated prior to the Early Medieval migrations of Early Slavs towards Central and South-Eastern Europe. Also, further studies are needed to discriminate between modern Polish lineages descending from the local West Baltic-speaking people (like Old Prussians and/or Yotvingians) and those representing some potential newcomers of East Baltic, mostly Lithuanian, origin.
The last question that has been investigated in our study is the presence of Ashkenazi Jewish-derived subclades in the modern Polish population. As a consequence of the Second World War, the Jewish population has been drastically reduced from nearly 10% in the prewar period to less than 0.1% in today’s Poland (Stankowski and Weiser 2011). However, in spite of strong religious, cultural and social barriers separating ethnic Poles from Ashkenazi Jews in historical times, the very long coexistence of these two populations could have resulted in significant contribution of Jewish Y-chromosomal lineages to the Polish Y-DNA pool. Our results suggest that this share is approximately 1.3%. Since the corresponding autosomal admixture is likely not evenly distributed among modern Poles, the median level of such admixture is probably much below 1%, thus in the range that makes reliable detection extremely difficult, if not impossible.

Comments (0)