
Remember me

Although both basal-like and triple negative breast cancers are associated with poor clinical outcomes [1], initially thought to be synonymous and moreover often using TNBC as a clinical surrogate term for BLBC repeatedly in the literature, they represent different biological phenomena [2]. TNBC is so named, based on clinical diagnosis, lacking three hormonal receptors- ER, PR and HER2 whereas BLBC is named based on molecular profiling characterized by the expression of basal-markers such as cytokeratins- KRT5/6, KRT17 which are expressed in outermost basal/myoepithelial cells of the normal breast milk ducts. TNBC is a highly heterogeneous subtype that has been divided into four refined molecular subtypes [3]. The basal-like subtype is the most predominant subtype within TNBCs. Therefore, there is a significant overlap between them, but not all TNBCs are basal-like breast cancer and likewise not all BLBCs are TNBCs [4,5,6]. According to the Lehmann et al., the four refined molecular subtypes are: basal-like 1 (BL1), basal-like 2 (BL2), mesenchymal (M), and luminal androgen receptor (LAR) based on distinct gene expression patterns [3]. Although several molecular subtypes, summarized chronologically in Fig. 1, have been proposed by different groups, the current consensus is that TNBC comprises of at least four major molecular subtypes and basal-like TNBC together comprises majority of the tumors. Each molecular subtype has its putative therapeutic targets, prognostic factors and predictive biomarkers based on the transcriptomic profiling and have different therapeutic vulnerability and response rates [7, 8]. While immunotherapies and other targeted therapies are still under development, chemotherapy remains the mainstay for BLBCs/TNBCs to date, due to the lack of targeted hormonal receptors in majority of these cancers [9]. Among the various intrinsic subtypes, basal-like TNBC, in particular basal-like 2 subtype exhibits stronger stem-like features, therefore potential strategy to address the clinical challenges of BLBCs is to target vulnerabilities in cellular plasticity [10,11,12].
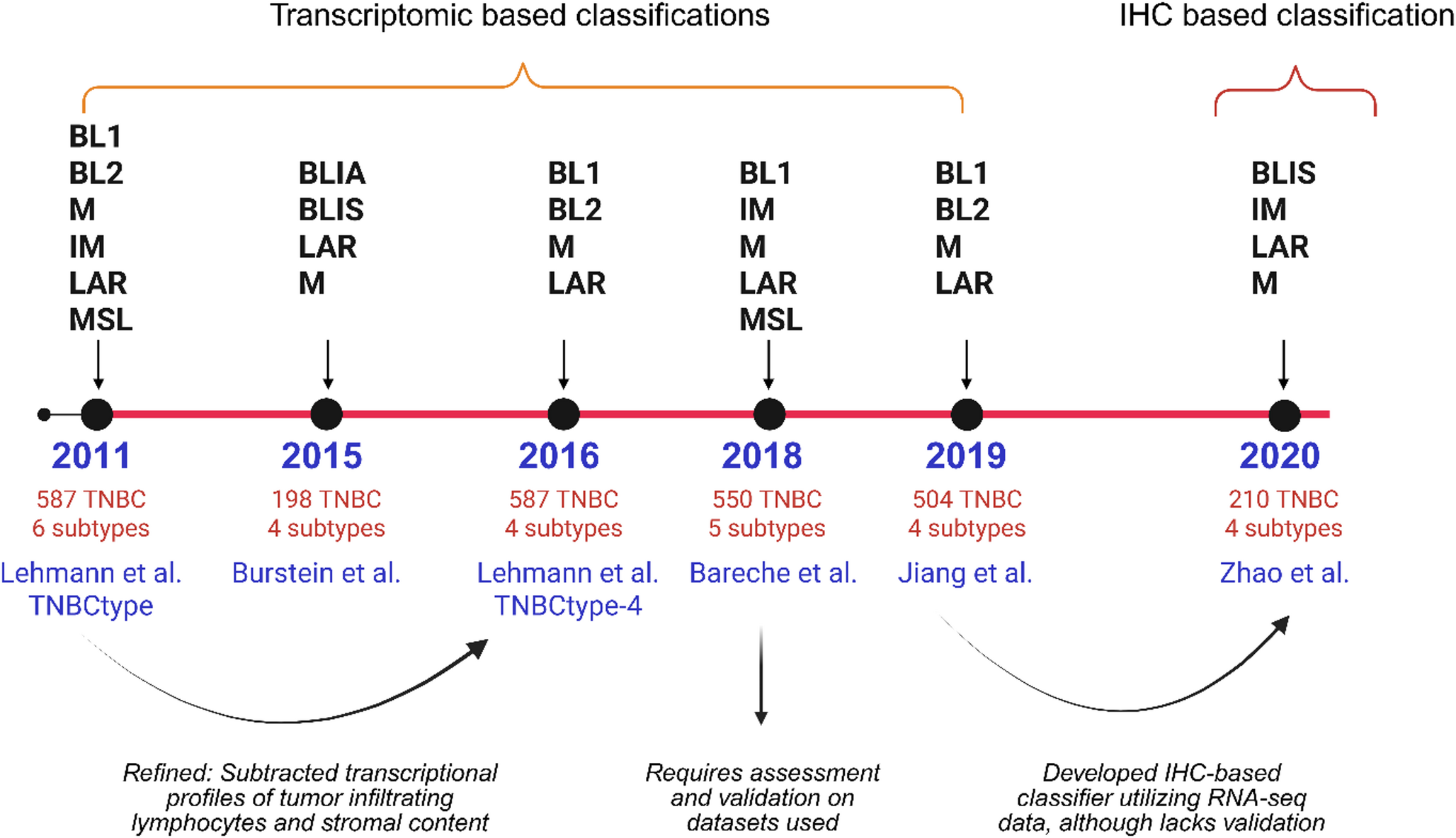
Fig. 1
TNBC subtype classifications in chronological order. Schematic illustration shows various classifications of the intrinsic molecular subtypes under TNBC year wise based on different groups [3, 13,14,15,16,17]. BL1: basal-like 1, BL2: basal-like 2, IM: immunomodulatory, LAR: luminal androgen receptor, MSL: mesenchymal stem-like, BLIA: basal-like immune active, BLIS: basal-like immune suppressed, M: mesenchymal
The strong link between BRCA1 mutation, basal-like characteristics, and BLBCs [18,19,20] raises the question of whether the loss of BRCA1 function contributes to the development of sporadic BLBCs, and whether this connection could be leveraged therapeutically to create new chemotherapeutic drugs and biological treatments.
At the molecular level, BRCA1 is best known for its DNA repair function, in maintaining genome integrity by resolving complex hybrid DNA: RNA or DNA: DNA structures such as R-loops [21, 22] and D-loops [23] respectively, along with other well-reported functions in the literature. Despite this, it is not clear whether these alterations of BRCA1 functions affect all cell types equally within breast tissue or if there is a preferential association of BRCA1 genome-wide among particular cell type over others, resulting in cell lineage-specific tumorigenesis in the breast epithelium of women carrying BRCA1 mutations. Additionally, an important question is whether BRCA1 loss of function could result in cellular or lineage plasticity. Lineage plasticity refers to the ability of a cell type to switch from one state to another, dynamically and reversibly, and is a major concern in cancer treatment options [24, 25] hence it would be meaningful to comprehensively summarize the current state of BRCA1 in terms of lineage plasticity. In this review, we shed light on the untapped role of BRCA1 in lineage plasticity. This perspective focuses on the new role of BRCA1 in promoting cellular plasticity, with an emphasis primarily limited to breast cancer.
The Cell of Origin of BRCA1-derived Breast TumorsBreast tissue is made up of a beautiful ductal tree network, which contains two compartments: luminal compartment in the interior side where luminal progenitors and mature luminal cells reside, and basal compartment coating the luminal layer where mammary stem cells and terminally differentiated myoepithelial cells called basal cells are located [26]. Each compartment has a number of stem-like cells called stem/progenitor cells, which maintain breast homeostasis by a continuous process of self-renewal and differentiation. Progenitors are partially differentiated stem-like cells considered downstream of real stem cells, having the properties to self-renew and differentiate into only specific/limited lineages [26]. To understand the mammary epithelial hierarchy, a single multipotent mammary stem cell (MaSC) serves as the origin of lineage-biased stem cells, basal stem cells and luminal stem cells [26, 27]. The basal stem cell population gives rise to committed myoepithelial progenitors, which subsequently differentiate into mature myoepithelial cells positioned in intimate association with the basement membrane. Luminal stem cells, on the other hand, generate a pool of long-term repopulating luminal stem cells with mixed lineage [28]. Previous studies have proposed that these mixed lineage luminal stem cells undergo bifurcation into two distinct luminal progenitor populations- ductal progenitors and alveolar progenitors each giving rise to terminally differentiated cell types: ER+/ER- ductal luminal cells and ER- alveolar luminal cells respectively [26]. Ducts, a network of thin tubes that carry milk from the lobules to the nipple, contain both ER- and ER + luminal cells, whereas alveolar luminal cells, lining the milk-producing alveoli are primarily ER- [26, 29].
Traditionally, it was thought that a common luminal stem/progenitor cell gives rise to all luminal cell types, encompassing both ER + and ER- populations [26]. However, the use of multiple approaches such as transplantation studies, flow cytometry studies, lineage tracing experiments and scRNA-seq to understand the mammary gland hierarchy, suggests this is not the case. Lineage tracing strategies allowed us to specifically label a particular cell type, in order to track the trajectories of the daughter cells during puberty and adult life. By now, it has been demonstrated that ER + and ER- luminal cells are maintained by their own pool of lineage-restricted stem cells using genetic lineage tracing experiments exploiting cell type-specific promoters. For example, studies using the ER-rtTA transgenic mice model in which only ER expressing cells were tracked throughout the development, showed that these ER + cells are maintained by their own lineage-restricted ER + stem cells [27]. Another study using Notch1-CreERT2SAT knock-in mice showed that NOTCH1 expressing progenitors specifically generate ER- luminal cells but not ER⁺ luminal cells [30]. Wang et al., using Sox9-CreERT2 and Prom1-CreERT2 mice delineated that ER + and ER- luminal cells are two independent lineages which are maintained by distinct progenitors, where ER + cells are maintained by PROM1 + expressing luminal progenitors whereas ER- cells are maintained by SOX9 expressing luminal progenitors providing a revised mammary epithelial cell hierarchy [31]. Figure 2 summarizes the most updated mammary epithelial hierarchy of stem and progenitor cells. Single-cell RNA sequencing (scRNA-seq) analyses have been applied to both human [32] and murine mammary tissues [33] to gain deeper insights of epithelial cell diversity and lineage relationships in an unbiased manner. This unbiased view of mammary gland hierarchy at a high cellular resolution largely supports the more accepted hypothesis of mammary gland hierarchy depicted in Fig. 2.
Fig. 2
Schematic updated model of mammary stem cells and progenitor cells hierarchy. Proposed model of the mammary epithelial cell differentiation hierarchy. A multipotent stem cell present during development gives rise to luminal epithelial and basal stem cell, which further divide into luminal and basal progenitors. ER- ductal and ER + alveolar luminal cells both arise from ER- luminal progenitor whereas ER + luminal progenitor makes only ER + luminal cells. The basal stem cell population gives rise to myoepithelial progenitors, which subsequently differentiate into mature myoepithelial cells
Breast cancers develop from either of the compartments—luminal or basal—and are primarily discussed as having either a luminal or basal origin accordingly. However, contrary to the earlier belief that specific subtypes arise from their respective progenitor cells [34]’[35], the majority of breast cancers originate from luminal cells, including the basal-like breast cancers, which surprisingly arise from the luminal progenitors [36,37,38,39]. This is due to the lineage plasticity which can be observed due to various genetic, epigenetic, and environmental factors which are never fixed determinants [40,41,42]. Genetic mutations can alter the structure of the epigenetic landscape, influencing the nature of lineage plasticity. In the context of genetic factors, loss-of-function mutations in TP53, RB1, PIK3KCA, and BRCA1 have been linked with lineage plasticity in multiple cancers [25, 43,44,45]. Although a considerable amount of literature talks about TP53 and/or RB1 in association with lineage plasticity among various cancers, to our knowledge, BRCA1 has never been summarized in terms of promoting lineage plasticity, especially in breast cancer. Of note, BRCA1 mutations increase the likelihood of developing cancers which are restricted mostly to the breast and ovary; why they do not affect other organs remains a mystery. Most of the tumors arising in patients with BRCA1 mutations are limited to a lethal, highly aggressive, and metastatic subtype, having basal-like phenotype [46, 47], raising a question of the basal stem cell as the origin of BRCA- derived breast tumors. Basal-like breast cancer generally carries a worst prognosis than other subtypes along with high rates of recurrence [8]. Reasons include but are not limited to the lack of the steroid hormone receptors (estrogen receptor and progesterone receptor) and human epidermal growth factor receptor 2, genomic instability and high rates of TP53 mutations [15], and a high proportion of breast cancer stem cells [48, 49].
Plasticity Induced by BRCA1 loss-of-functionCellular plasticity is a hotspot area in the horizon of cancer biology since it is one of the reasons for tumor recurrence and treatment failures. As new therapies are developed in the context of lineage plasticity, the goal of treatment may be to prevent, reverse, or treat the new lineage especially if the new lineage is worse than the original lineage [24]. Although most widely studied gene in the context of DNA-repair, BRCA1 is only moderately studied in association with cellular plasticity in breast cancer after TP53 and RB1 [36, 38, 50, 51] probably due to significant longer time taken by the tumors to form in the currently available BRCA1 tumor mouse models in research. Given the fact that most of the tumors arising in patients with germline BRCA1 mutations have a basal-like phenotype [47, 52], it was postulated that BRCA1 mutations in basal stem cells could be the driver for the basal-like breast cancer [53]. However, the origin of BRCA1-deficient breast tumors was questioned in later years.
Using discrete epithelial subpopulations derived from the breast tissues of normal and BRCA1 mutation carriers, it was demonstrated that the loss of BRCA1 is associated with aberrant amplification of luminal progenitors, which resembled the gene expression profiles of basal-like breast cancers rather than any other subtype [37]. Lim et al. identified that the fraction of luminal progenitors was much larger than basal subset (MaSC-enriched) in histologically normal mammary tissue obtained from BRCA1 mutation carriers with higher clonogenicity potential in these luminal progenitors compared to MaSC-enriched basal subset. Consistent with this, higher clonogenic activity was also reflected in the luminal progenitors isolated from mouse mammary tumor virus MMTV-Cre; Brca1f/f glands. Comparative unbiased gene expression profiling of the four mammary cell subpopulations- MaSC-enriched, luminal progenitor, mature luminal epithelial and stromal with the various breast cancer subtypes from human normal breast tissue revealed the closest association of basal breast cancers with luminal progenitors. Additionally, the signature expression score of the luminal progenitors was indeed highest in basal subtype of breast cancer indicating a clear correlated expression between these two [37]. To explore the relationship between BRCA1 dysfunction and the basal phenotype, researchers established mammary gland specific- Brca1 conditional knockout mouse models. One such mouse model is Blg-Cre; Brca1f/f; p53+/−, which harbors a conditional Brca1 loss-of-function in the luminal progenitor compartment, while Blg promoter is active in almost all progenitor cells in the luminal estrogen receptor negative population. To understand the genetics of this mouse model properly, Brca1 floxed homozygous mice were crossed with p53-deficient mice forming Brca1f/fp53+/− mice line. The resultant littermates were further crossed with Blg-Cre transgenic mice to generate mice homozygous for the floxed allele of Brca1, heterozygous for p53, and carrying the Cre transgene [54]. Targeted expression of Cre recombinase in luminal progenitors resulted in the deletion of Brca1 leading the formation of mammary tumors.
Another mouse model is K14-Cre; Brca1f/f; p53+/− mice, in which Cre expression is driven specifically in the basal compartment of the mouse mammary epithelium since Keratin 14 (K14) is expressed in basal cells. Although various groups [36, 53, 55] have reported different observations utilizing a mouse model of basal cell specific K14-Cre-driven BRCA1 loss, the tumor spectrum they observed was similar, exhibiting features characteristic of human BRCA1-mutated basal-like breast cancer. The reason behind these discrepancies could be attributed to the status of p53, where one group used p53+/− whereas another used p53−/− to initiate the malignant transformation. This selection pressure imparted by the status of p53 could alter the relative likelihood of targeting Brca1 loss in stem versus progenitor cells [56], resulting in tumor formation from luminal progenitors in K14 model [53]. Molyneux et al. used the two models described above to understand the origin of human sporadic basal-like breast cancers. Lineage-specific BRCA1 knockout in luminal progenitors and basal stem cells using Blg-Cre; Brca1f/f; p53+/− and K14-Cre; Brca1f/f; p53+/− mouse models respectively demonstrated that only tumors originating from BRCA1 KO luminal progenitors resemble the features similar to those of human basal-like breast cancers with a similar histological appearance whereas the tumors arising from BRCA1 KO basal stem cells neither resembled human BRCA1 nor sporadic basal-like breast cancers, other than just having the expression of basal-markers [36].
Using a “humanized” model system, Proia et al. further confirmed that BRCA1mut/+ cells give rise to basal-like breast tumors, whereas BRCA1+/+ cells generate both luminal and basal-like tumors [57], supporting the previous observation of a probable link between BRCA1 mutation and the “cell of origin” for basal-like breast cancers. The group examined the biology of disease-free breast tissues from patients harboring deleterious mutations in BRCA1. They generated human breast tumors in vivo by first introducing a set of oncogenes (P53R175H, CCND1, PI3KCA, RasG12V- reflective of both the luminal and basal tumor categories, to remove the lineage specific bias) into dissociated single epithelial cell suspensions obtained from reduction mammoplasty tissues and then reintroducing the suspension into the humanized mammary fat pad. Characterization of the tumors, both phenotypically and genomically, developed by BRCA1+/+ and BRCA1mut/+ suspensions revealed that luminal progenitors from BRCA1mut/+ tissue had attenuation of gene expression associated with luminal differentiation and retained features of basal differentiation, and the basal phenotype of BRCA1-associated tumors resulted from the pre-existing increased basal differentiation state of the luminal progenitor population.
Although the basal-like breast cancer-promoting effect of BRCA1 has been described above, how BRCA1 mutation affects the luminal progenitor cell state remains poorly understood. To capture early impact of genetic aberrations on cell types during premalignant transformation, upon loss of Brca1, time-resolved single-cell profiling was performed on the cells isolated from the mammary glands spanning various premalignant stages using Blg-Cre; Brca1f/f; p53+/− TNBC mouse model [38]. The only cluster that was seen to expand was a cluster of luminal cells. Furthermore, chromatin accessibility via ATAC-seq showed increased accessibility scores around key transcription factors such as Cebpb, Elf5 [58], Nfkb1 [59, 60] and Sox10 [61, 62], and the majority of them are linked with basal-like breast cancer. Analysis of both mouse Blg-Cre; Brca1f/f; p53+/− and the human BRCA1-mutant epithelial cells proved that BRCA1 loss led the aberrant expansion of lineage progenitors. The study highlighted the fact that altering Brca1/p53 status promotes a differentiation program hardwired in luminal progenitors [38]. All these findings were in support of another similar study published in the same year utilizing single cell RNA sequencing approach further confirming the relevance of luminal cells as the potential originator of breast cancers and that luminal progenitors are the probable cells of origin for basal-like breast cancers [63]. Altogether, these studies shed light on the important role of BRCA1 in promoting luminal-to-basal lineage transdifferentiation.
Mechanisms by which BRCA1 Loss of Function Promotes PlasticityBy now, it is well established that basal-like breast cancer most likely originates from luminal progenitors with BRCA1 mutation/loss-of-function; what remains opaque is how these genetic aberrations drive cellular plasticity. Does BRCA1 act by controlling key plasticity factors, such as members of the SOX family (SOX9 [64], SOX10 [65], SOX2 [66]) or SLUG [57, 67, 68] or other transcription factors such as ONECUT2 [69]? These transcription factors have been well documented as promoting lineage plasticity via lineage tracing experiments. Several lines of evidence suggest underlying possibilities. One possibility is via BRCA1 regulation of SLUG, a basal-lineage-biased transcription factor which is regulated indirectly by BRCA1 [68]. BRCA1 appears to regulate SLUG protein stability, however, the precise mechanism of BRCA1 mediated SLUG stability remains obscure [57]. SLUG is expressed primarily in basal-stem-cell-enriched population in both mice and humans [70]. Elevated expression of SLUG mRNA is seen in breast cancer cell lines having basal-like breast carcinoma phenotype. Moreover, overexpression of SLUG in luminal type MCF-7 cells transformed them into a basal-like phenotype, accompanied by the acquisition of gene expression pattern similar to that of basal-like breast cancer [71].
Another potential mechanism may involve its DNA-regulatory function. BRCA1 has been shown to regulate lineage plasticity through its normal DNA damage repair mechanism [72, 73] and dysregulated expression of BRCA1 contributes to the loss of lineage integrity, which underlies tumor heterogeneity, as modelled and depicted in Fig. 3. Induced loss of BRCA1 in vivo has been shown to induce luminal-to-basal transdifferentiation; most probable reason is the insufficient availability of DNA repair proteins, which includes BRCA1 itself and its binding partners, NUMB and HEY1 [73]. Targeted ablation of Brca1 in the mammary gland’s luminal cells, achieved by intraductal injection of the Cre-expressing adenovirus under the Krt8 promoter, led to deletion of floxed Brca1 and/or Trp53 knockout, resulting in aberrant transdifferentiation of luminal cells causing basal-like breast tumorigenesis. Numb knockout mammary epithelial cells, when grown in vitro form hyperproliferative structures in contrast to the hollow-type acinar structures characteristic of in vitro-grown basal-like cells and promotes tumorigenesis which is p53 dependent [74]. These effects are as similar as to the Brca1 depleted phenotype suggesting that NUMB might, in part, play a role in cell fate determination via BRCA1-NUMB proteins complex in the nucleus. Luminal progenitors are highly proliferative which indeed call for hi
Comments (0)