
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Remember me
The brain exhibits continuous intrinsic functional activity, even in the absence of external stimuli [1, 2]. This spontaneous activity is hierarchically organized across multiple spatial scales, ranging from neural connections within a single region to local networks, which integrate into large-scale networks and global brain states [3–5]. Understanding the causal effects of targeted local perturbations across this hierarchy up to global brain dynamics is essential for elucidating principles of neural information flow and improving the efficacy of therapeutic brain stimulation [6]. However, the mechanisms through which focal perturbations to a single brain region can immediately influence global brain dynamics remain unclear. There are several reasons why investigating this question has been a challenge. First, ongoing brain activity organizes into discrete, recurring global configurations that remain quasi-stable for tens of milliseconds before transitioning to alternative configurations [7–9]. Characterizing how perturbations influence these rapidly evolving states requires millisecond-level temporal resolution to detect which configurations emerge following stimulation. Current connectivity-based approaches examine how stimulation alters functional coupling between brain region pairs [10, 11] but cannot capture reconfigurations of global brain states. Second, focal perturbations induce both time-locked and non-time-locked activity relative to stimulation onset [12, 13]. While trial averaging captures time-locked responses, it cancels out non-phase-locked induced activity that varies across trials. Most existing methods rely on trial averaging [14, 15], preventing characterization of how perturbations influence global brain states at the single-trial level. Accordingly, it is not well understood how focal changes in the ongoing activity of a single brain region can rapidly influence the dynamics of global brain states. Insight into these mechanisms is key to furthering our understanding of how information flows between different spatial scales and how this relates to brain functioning in health and disease.
A promising framework for investigating the neurophysiological effects of focal perturbations with high temporal resolution is the simultaneous combination of transcranial magnetic stimulation (TMS) and electroencephalography (EEG) [16–18]. TMS-EEG studies have shown that single pulses of TMS can perturb ongoing electrophysiological activity at the stimulation site by inducing changes to the local oscillation phase dynamics [19–22], the effects of which then propagate and evoke activity in distal regions [23, 24]. The extent and pattern of this downstream propagation depend on which specific brain region is targeted [25]. However, much of our current understanding of TMS-evoked potentials (TEPs) derives primarily from EEG analyses characterizing peak amplitudes and latencies at selected electrodes or whole-scalp metrics such as global field power [26]. While amplitude-based approaches can be used to quantify the magnitude and topographic distribution of perturbation effects, they do not directly assess effects on the functional connectivity or global brain states and their dynamics. Connectivity-based methodologies have been developed to quantify inter-regional coupling dynamics following TMS perturbations [27–30]. For instance, directed transfer function analysis applied to task-locked TMS-EEG data during saccadic eye movements has quantified frequency-specific information flow between electrode sites [31]. This approach revealed site-specific perturbation effects whereby prefrontal cortex stimulation modulated fronto-occipital connectivity patterns immediately following target onset, whereas posterior parietal cortex stimulation elicited delayed connectivity changes in parieto-temporal pathways. Moreover, graph theoretical analyses have demonstrated that focal TMS perturbations can systematically modulate trial-averaged sensor-level functional connectivity patterns [32]. These findings collectively suggest that focal cortical perturbations can transiently reconfigure regional connectivity to facilitate information processing between brain regions. While these methods provide insights into regional interactions, sensor-level connectivity estimates are influenced by volume conduction [33, 34]. Moreover, decomposing effects into pairwise relationships does not directly capture the global brain states and their dynamics. Whether focal perturbation triggers reorganization of global brain states remains unexplored, requiring a shift from regional or pairwise descriptions toward representation of the brain’s moment-to-moment global state at the single-trial level.
EEG microstate analysis provides a complementary analytical framework that addresses several of these limitations by representing whole-brain activity as a sequence of discrete global states rather than decomposed regional or pairwise relationships [35, 36]. In this approach, the entire scalp topography at each time point is assigned to a single canonical state class, thereby preserving the multi-channel configuration as an integrated unit [37]. Resting-state EEG studies have consistently identified four to seven canonical microstate classes. Although not completely equivalent, each microstate class has been associated with activation patterns overlapping the known functional magnetic resonance imaging (fMRI)-based large-scale functional networks. Transitions between microstate classes may therefore reflect rapid reconfigurations of large-scale network dynamics [38, 39]. Importantly, microstate analysis operates on reference-independent topographic patterns, which reduces sensitivity to volume conduction effects relative to electrode-wise amplitude or connectivity measures. Recent work has extended microstate analysis to TMS-evoked responses, demonstrating that the post-pulse microstate sequence following primary motor cortex stimulation varies systematically with pulse waveform and electric-field direction in trial-averaged data [40]. However, two critical questions remain to be answered. First, whether focal TMS influences microstate dynamics at the single-trial level, where moment-to-moment variability in brain states may interact with perturbation effects, has not been established. Second, whether these perturbation-induced alterations exhibit site-specificity across functionally distinct cortical regions remains unknown. The millisecond temporal resolution of microstate analysis is well-suited for addressing these questions. This can be achieved by examining whether single TMS pulses systematically modulate the occurrence probability of specific microstate classes at each post-pulse time point relative to baseline, and whether these effects differ across stimulation sites in a manner consistent with the functional roles of the targeted brain regions.
In this study, we aimed to understand how TMS applied to specific cortical regions can affect global brain state dynamics as measured by EEG microstates. We analyzed data from healthy participants at rest who received single-pulse TMS-EEG over three cortical sites: the left and right dorsolateral prefrontal cortex (DLPFC) and the left primary motor cortex (M1). These regions were chosen due to their association with distinct large-scale functional networks, as the DLPFC is associated with the frontoparietal network and executive functioning, and the M1 with the primary motor network and motor functioning [41, 42]. Given that EEG microstates capture global-scale functional activity of specific brain networks [43, 44], and that TMS-evoked responses vary significantly across cortical regions [45–47], we hypothesized that these localized perturbations would generate site-specific microstate reorganization, with bilateral DLPFC stimulation inducing significantly different microstate occurrences and transitions compared to M1 stimulation. We expected TMS to have the greatest effects on the dynamics of microstates that have been linked to the functional processes of the targeted region; specifically, microstate D, which has been consistently associated with executive functioning [44], would increase following DLPFC stimulation, and microstate A, which has been linked to low-level sensory processing areas and sensory input processing during motor preparation and motor task performance [48, 49], would increase following M1 stimulation.
2.1. ParticipantsThirty-six healthy individuals (13 males; mean age ± SD: 21.1 ± 2.0 years) with no history of psychiatric or neurological conditions were recruited at the Centre for Addiction and Mental Health in Toronto, Ontario, Canada. The study protocol was approved by the Centre for Addiction and Mental Health in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants, and a screening process was conducted prior to enrollment to ensure their suitability for receiving TMS.
The generalizability of findings from dataset 1 was assessed through analysis of an independent, publicly available TMS-EEG dataset (dataset 2) in which two of the three brain regions (right DLPFC and left M1) were examined as in dataset 1 [46]. Dataset 2 was comprised of healthy participants (n = 20, 9 males; mean age ± SD: 24.3 ± 3.1 years), from whom informed consent was obtained under a protocol approved by the NIH Combined Central Nervous System Institutional Review Board. All ethical guidelines established in the Declaration of Helsinki were adhered to in this supplementary investigation.
2.2. TMS-EEG procedureDataset 1
T1-weighted MRI was performed for each participant to facilitate precise TMS coil positioning during experimental sessions. Neuro-navigation (Brainsight TMS Navigation; Rogue Resolutions) was employed for accurate targeting of specific cortical regions of interest. Resting motor threshold (rMT) for each hemisphere was determined and defined as the minimum stimulator output required to elicit a motor-evoked potential (MEP) exceeding 50 μV in at least five out of ten trials [50]. Stimulator intensity was subsequently calibrated to 120% of the rMT, to generate a 1 mV MEP across 15 consecutive trials. When the average MEP fell outside the 0.5–1.5 mV range, adjustments to stimulator intensity were implemented until the desired response amplitude was achieved.
A series of 80 monophasic TMS pulses was administered to each of the three stimulation sites separately (left DLPFC (80 pulses), right DLPFC (80 pulses), and left M1 (80 pulses)) with a consistent 5 s interstimulus interval. Stimulation was delivered via two Magstim 200 stimulators (Magstim Company) equipped with a 70 mm figure-of-8 coil. For DLPFC targeting, MNI coordinates (−35, 45, 38) and (35, 45, 38) were utilized for left and right hemispheres, respectively [51, 52]. Optimal coil location and orientation for motor cortex stimulation were determined by sites where maximal MEPs were elicited, as recorded via electromyography (EMG) from the abductor pollicis brevis muscle. A thin foam layer was positioned between the TMS coil and the participant’s scalp to minimize electrode movement and attenuate auditory TMS-EEG artifacts caused by bone conduction [53]. Earplugs were provided throughout the session to further reduce auditory-evoked potentials associated with TMS discharge [53, 54]. During the TMS sessions, participants were instructed to keep their eyes open and focus on a black cross in front of them.
Twenty-four participants completed a follow-up TMS-EEG session approximately two weeks after initial testing. All three stimulation sites (left DLPFC, right DLPFC, left M1) were targeted using identical procedures, enabling within-subject assessment of result reproducibility. This approach allowed us to determine whether the region-specific microstate dynamics observed in the primary analysis could be consistently replicated in the same participants when tested at a separate time point. Comprehensive reproducibility analyses demonstrating consistency of the observed effects across independent sessions are presented in the supplementary materials.
Sham-controlled stimulation was also conducted with 25 participants from dataset 1. Participants were randomly assigned to receive sham stimulation over either the left DLPFC (n = 12) or right DLPFC (n = 13), but not both hemispheres. Each participant received 80 single pulses to their assigned sham site, matching the pulse count used in active stimulation conditions. The TMS coil was rotated 90° during sham conditions, preserving characteristic sensory inputs while minimizing magnetic field penetration into cortical tissue. Statistical comparisons between active and sham conditions were conducted by pooling data from left DLPFC and right DLPFC stimulation sites; these active-sham analyses are detailed in the supplementary materials.
Dataset 2
Neuro-navigation (Brainsight frameless system) was employed for targeting left M1 and right DLPFC. Monophasic TMS pulses were delivered via a 70 mm figure-of-eight coil connected to a Magstim 200 stimulator, while EMG activity was recorded from the right first dorsal interosseous muscle using bipolar surface electrodes. The optimal M1 scalp location was identified as the site where minimal stimulation intensity was required to elicit at least five MEPs >50 µV across ten consecutive trials. rMT was determined using an automated threshold-tracking algorithm. Stimulation intensity was calibrated to each participant’s rMT (45.9 ± 9.37% [mean ± SD] of maximum stimulator output). The experimental protocol consisted of 150 active TMS pulses delivered to each stimulation site in counterbalanced order at 5 s intervals. A 15% temporal jitter was incorporated to minimize anticipatory effects. Foam earplugs were inserted during all TMS procedures to attenuate auditory artifacts. Additional methodological details can be found in the corresponding manuscript [46].
2.3. EEG recording and preprocessingIn dataset 1, EEG data were acquired using a 64-channel Synamps 2 system (Compumedics Neuroscan) with electrodes referenced to a site posterior to Cz and data captured in DC mode at 20 kHz. Dataset 2 utilized a TMS-compatible BrainAmp MR+ system (Brain Vision) to sample EEG signals at 5000 Hz (DC filtered at 1000 Hz) from 30 scalp electrodes arranged in the 10–20 system, with electrode impedances maintained below 10 kΩ. A semi-automated preprocessing pipeline was applied to all EEG recordings from both datasets following current best practices for TMS-EEG artifact removal (detailed procedures are provided in the supplementary materials).
2.4. Microstate analysisMicrostate analyses were conducted on cleaned and epoched EEG data from all experimental conditions, with dataset 1 and dataset 2 analyzed separately due to different EEG coordinates. Data were bandpass filtered (1–40 Hz) and analyzed using a modified k-means clustering approach to identify five prototypical microstate topographies [55]. Topographic maps were extracted by detecting local peaks in global field power, with temporal labeling performed using standard smoothing algorithms (see supplementary materials for detailed parameters).
2.5. Characterization of TMS-induced microstate dynamicsFollowing microstate identification, canonical topographies were back-fitted, and each time-point was assigned a microstate label based on spatial similarity to the predefined maps. This procedure yielded trial-wise microstate sequences centered on a single TMS pulse and spanning one second before to one second after stimulation. For analysis, two intervals were contrasted: a baseline period (−1000 to −10 ms) and a post-pulse period (+20 to +1000 ms), with the −10 to +20 ms window excluded to minimize residual stimulation artifacts.
Relative occurrence frequency (ROF)
ROF quantified how often each microstate appeared following TMS relative to baseline. At each time point, we calculated the proportion of trials exhibiting each microstate, creating temporal profiles of microstate prevalence. To account for the compositional nature of these proportions (which necessarily sum to one), we applied a centered log-ratio transformation, enabling meaningful comparisons on a ratio scale [56]:
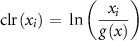
where 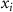 is the proportion of microstate
is the proportion of microstate 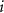 and
and 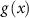 is the geometric mean across all microstate proportions at that time point. To handle time points where specific microstates were unobserved, a multiplicative replacement method was applied before transformation [57]. Zero values were replaced with
is the geometric mean across all microstate proportions at that time point. To handle time points where specific microstates were unobserved, a multiplicative replacement method was applied before transformation [57]. Zero values were replaced with 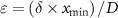 , where
, where 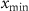 is the smallest non-zero proportion,
is the smallest non-zero proportion, 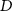 is the number of microstate classes, and
is the number of microstate classes, and 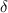 is a scaling factor set to 0.5 to ensure a small replacement value. Non-zero components were adjusted proportionally to preserve the compositional constraint. Each microstate’s ROF time course was baseline-corrected by subtracting, from all post-pulse time points, the median baseline ROF for that microstate computed over −1000 to −10 ms. The resulting ROF represents log-fold changes from baseline: positive values indicate increased microstate prevalence after TMS, while negative values indicate decreased prevalence. This metric thus captures how TMS redistributes the temporal coverage of distinct brain states.
is a scaling factor set to 0.5 to ensure a small replacement value. Non-zero components were adjusted proportionally to preserve the compositional constraint. Each microstate’s ROF time course was baseline-corrected by subtracting, from all post-pulse time points, the median baseline ROF for that microstate computed over −1000 to −10 ms. The resulting ROF represents log-fold changes from baseline: positive values indicate increased microstate prevalence after TMS, while negative values indicate decreased prevalence. This metric thus captures how TMS redistributes the temporal coverage of distinct brain states.
Relative transition frequency (RTF)
RTF measured how TMS altered the directional flow between microstates. We identified all transitions from one microstate to another (excluding self-transitions) and calculated their frequency at each time point, normalized by trial count. Analysis focused on the 20–500 ms post-pulse window, where stimulation effects are neurophysiologically expected [10]. Transition frequencies within this window were averaged and baseline-corrected against pre-pulse values (−1000 to −10 ms). Positive RTF values indicate transitions that became more frequent after TMS, while negative values indicate suppressed transitions. Critically, RTF is directional (i.e. transitions from state A to B differ from B to A) revealing asymmetric reorganization patterns in network dynamics beyond what state occupancy alone can capture.
2.6. Statistical analysisTMS effects on microstate occurrence frequencies
One-sample t-tests were conducted on the ROF of each microstate at every time point from +20 to +1000 ms relative to TMS pulse onset, testing the null hypothesis that ROF would not differ from zero following TMS. To control for multiple comparisons across time points, we employed threshold-free cluster enhancement (TFCE) with sign-flipping permutation testing (5000 permutations, p < 0.05) [58–60]. This approach preserves temporal dependence and controls family-wise error without an arbitrary cluster-forming threshold. Effect sizes for significant clusters were quantified using Cohen’s d, with the average across all significant clusters reported as the summary measure [61, 62]. Detailed TFCE methodology is provided in supplementary materials.
TMS effects on microstate transitions
Changes in transition frequencies between microstate pairs were evaluated using one-sample t-tests on the RTF for each pair during the post-pulse window (20–500 ms), testing against zero mean. Bonferroni correction was applied across the 20 microstate pairs (p < 0.05). Cohen’s d was calculated for each significant finding.
Site-specificity analysis
Site specificity of TMS effects was assessed separately for microstates’ ROF and RTF between stimulation sites. For ROF analysis, sites were contrasted through application of TFCE with sign-flipping permutation testing to the subject-level difference vector across the +20 to +1000 ms interval relative to pulse onset (5000 permutations; two-sided p < 0.05). In the results section, site contrasts exhibiting significant ROF differences are reported together with their direction and summary effect size (average Cohen’s d across significant clusters). Full TFCE parameters, cluster extents and peaks, exact p-values, and confidence intervals are provided in the supplementary materials.
For RTF analysis, site differences in baseline-corrected transition frequencies were tested for each of the 20 microstate pairs within 20–500 ms against zero, with family-wise error rate controlled across pairs via Bonferroni correction (two-sided p < 0.05/20). In the results section, only those transition pairs surviving correction are reported, along with their direction and Cohen’s d values. Detailed pairwise statistics (t-values, corrected p-values, confidence intervals) and multiple-comparison procedures are presented in the supplementary materials.
Replicability analysis
The robustness and generalizability of findings were assessed through replication and validation protocols. First, the reproducibility of results was examined in a subset of participants (n = 24) who underwent a second experimental session with identical stimulation parameters at left and right DLPFC and left M1 sites. The same preprocessing pipeline and statistical framework were applied to determine whether the same significant effects could be reproduced in this independent session. Second, the generalizability of effects was tested in an independent cohort (n = 20) in which right DLPFC and left M1 were stimulated using the same experimental protocol. The identical analytical approach was systematically implemented to determine whether the observed patterns could be reproduced across different participant samples. Whether the key ROF and RTF findings were directionally consistent and statistically significant under the same correction criteria across both validation datasets is reported in the results section. Detailed comparisons of effect sizes, correlation analyses between sessions, and comprehensive statistical outputs for both replication studies are presented in the supplementary materials, along with a discussion of any discrepancies or variations observed between the original and validation analyses. The reliability of TMS effects on RTF between microstate pairs was specifically evaluated through correlation analyses of Cohen’s d scores across repeated sessions and the independent dataset (presented in supplementary materials), examining both within-site reproducibility and cross-regional relationships.
All statistical analyses were performed using MATLAB R2024a (MathWorks Inc., Natick, MA). Custom scripts were developed for data analysis, and cluster-based permutation tests were implemented following established methods in the literature [63].
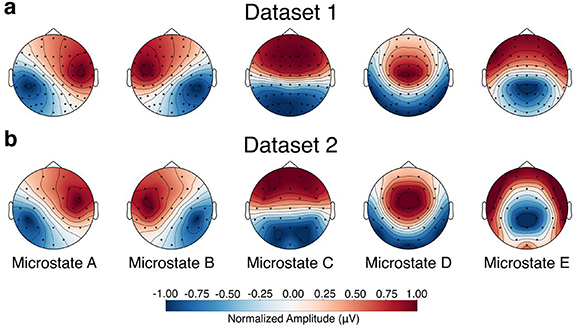
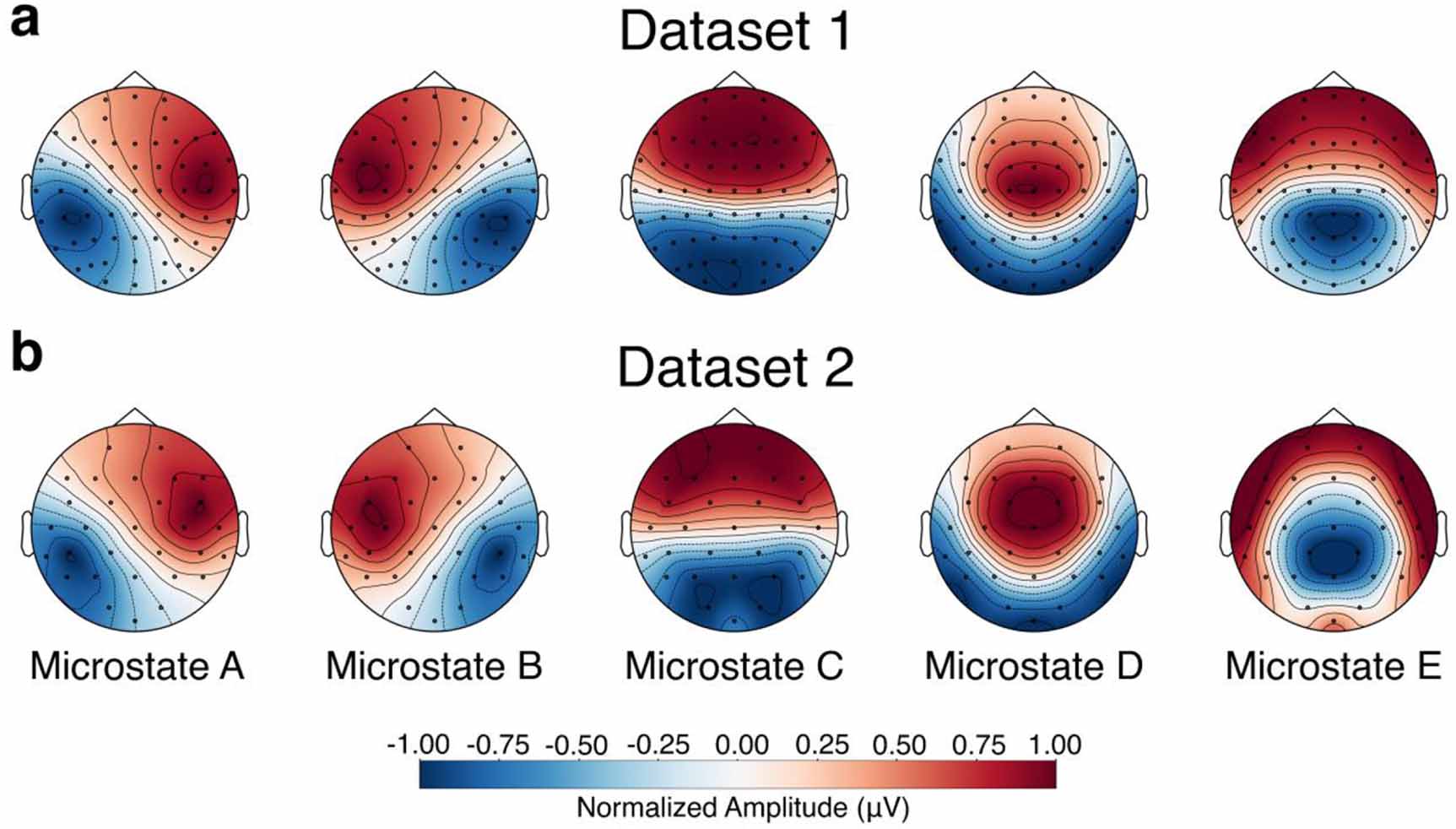
3.1. Dynamics of EEG microstates before and after TMS interventionIn both dataset 1 and dataset 2, we identified five canonical microstates that explained approximately 70% of the total variance. As illustrated in figure 1, the spatial topographies of the extracted microstates closely resembled those commonly reported in the literature and were consistent across both datasets. These microstates were labeled according to their topographical appearance as A, B, C, D, and E, in accordance with recent recommendations [44, 64]. We then backfitted these microstates to the original datasets for analysis of their dynamics.
Figure 1. Topographic maps of the extracted microstates. (a) Microstate topographies for dataset 1, where data from all stimulation sites and sessions were combined to generate a consistent set of maps. (b) Microstate topographies for dataset 2. Microstates in both datasets were labeled according to standard conventions in the field.
Download figure:
Standard image High-resolution imageTo investigate TMS-induced changes in microstate dynamics, we analyzed EEG data recorded one second before and after each pulse. For each subject, we quantified microstate changes using two measures: ROF, which calculates how often each microstate appeared post-pulse compared to its pre-pulse baseline across all trials, and RTF, which quantifies how the transition probability between specific microstate pairs changed after stimulation by comparing the averaged post-pulse transition rates to their corresponding pre-pulse baseline averages across all trials.
3.2. Changes in EEG microstate dynamics following TMSDataset 1—left DLPFC
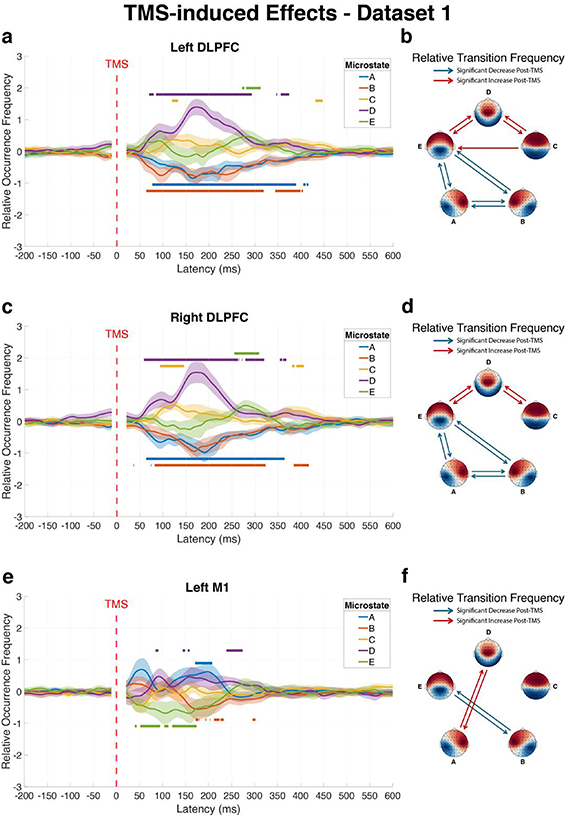
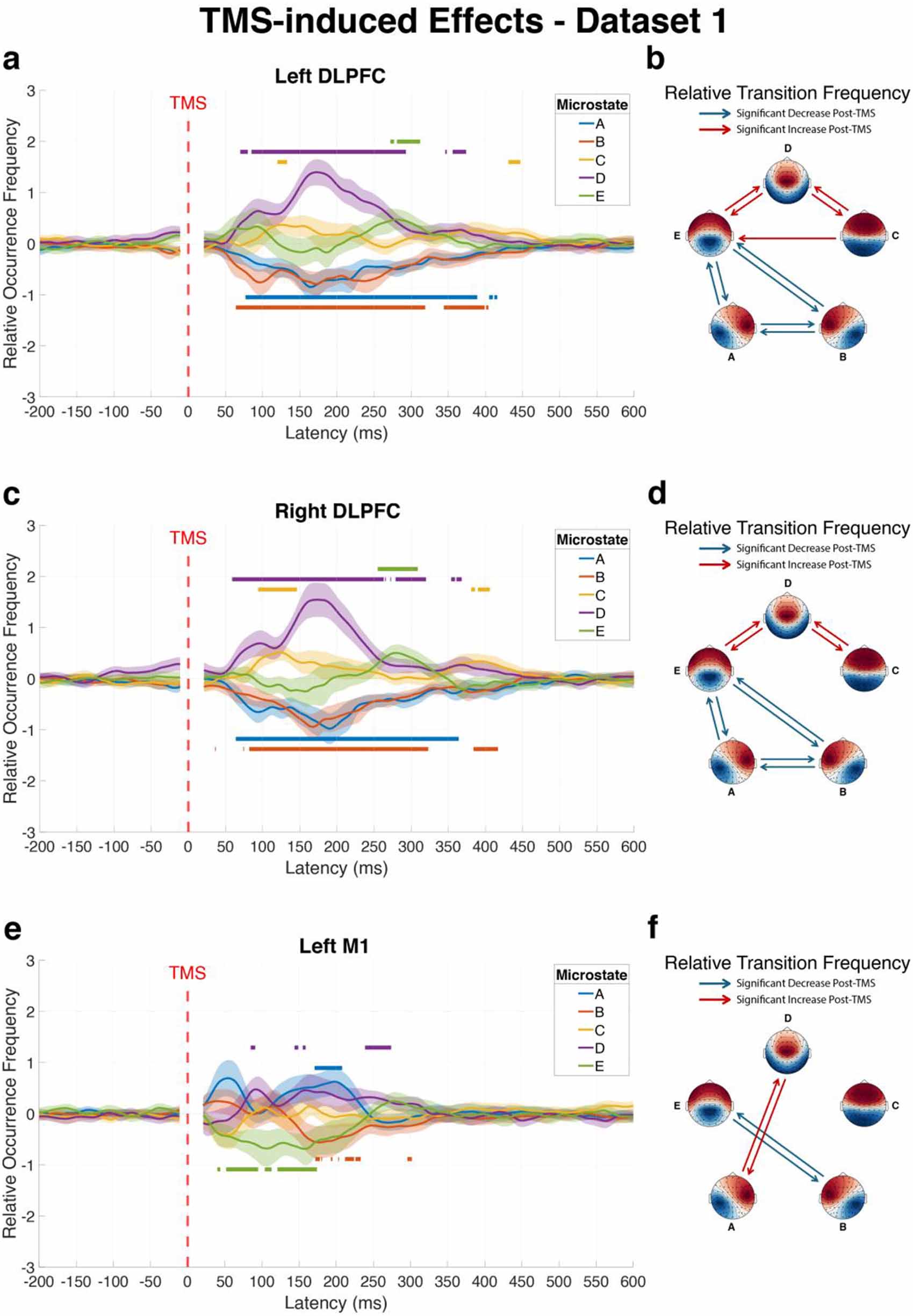
Single-pulse stimulation of the left DLPFC significantly altered the dynamics of all five EEG microstates within the first 500 ms after pulse onset (figure 2(a)). Analysis using TFCE (p < 0.05, 5,000 permutations) revealed distinct temporal patterns in the ROF of microstates. Following stimulation, microstates A and B showed significant decreases in ROF (Cohen’s d = −0.74 and −0.84, respectively), while microstates C, D, and E exhibited significant increases (Cohen’s d = 0.69, 0.95, and 0.72, respectively).
Figure 2. Significant alterations in microstate dynamics following TMS to different cortical targets compared to baseline (dataset 1, visit 1). The figure presents changes in brain microstates following TMS to left DLPFC (a) and (b), right DLPFC (c) and (d), and left M1 (e) and (f). Left panels (a), (c) and (e) show the average ROF trajectories of each microstate over time across subjects, with shaded areas representing 95% confidence intervals. Horizontal lines above or below curves indicate time intervals with statistically significant increases or decreases relative to baseline, respectively. Right panels (b), (d) and (f) display matrices of significant changes in RTF between microstate pairs during the post-pulse window (20–500 ms) compared to baseline, where red connections indicate significantly increased transitions and blue connections represent significantly decreased transitions.
Download figure:
Standard image High-resolution imageThe RTF between microstate pairs was also significantly altered following left DLPFC stimulation (figure 2(b)). Using Bonferroni correction for multiple comparisons (adjusted p < 0.05), we identified the transitions between microstates C, D, and E to have significantly increased: C–D (p < 0.001, Cohen’s d = 1.52), C-E (p = 0.039, Cohen’s d = 0.57), D–C (p < 0.001, Cohen’s d = 1.80), D–E (p < 0.001, Cohen’s d = 1.10), and E–D (p < 0.001, Cohen’s d = 0.88). In contrast, transitions involving microstates A and B significantly decreased: A–B (p < 0.001, Cohen’s d = − 1.90), A–E (p < 0.001, Cohen’s d = −1.04), B–A (p < 0.001, Cohen’s d = −1.65), B–E (p < 0.001, Cohen’s d = −1.01), E–A (p < 0.001, Cohen’s d = −0.81), and E–B (p < 0.001, Cohen’s d = −0.80).
Dataset 1—right DLPFC
Application of TMS to the right DLPFC also induced significant alterations in EEG microstate patterns (figure 2(c)). Within the first 500 ms after pulse onset, all five microstates showed significant changes in ROF. Microstates A (Cohen’s d = −1.51) and B (Cohen’s d = −0.77) showed significant decreases in ROF following the pulse. In contrast, Microstates C (Cohen’s d = 0.81), D (Cohen’s d = 0.67), and E (Cohen’s d = 0.89) exhibited significant increases in ROF.
The RTF between microstates showed significant alterations following right DLPFC stimulation compared to baseline (figure 2(d)). Specifically, the transition pairs C–D (p < 0.001, Cohen’s d = 1.31), D–C (p < 0.001, Cohen’s d = 1.44), D–E (p < 0.001, Cohen’s d = 1.14), and E–D (p < 0.001, Cohen’s d = 0.98) showed significant increases following the stimulation. In contrast, significant RTF decrease was found for transitions A–B (p < 0.001, Cohen’s d = −2.15), A–C (p = 0.009, Cohen’s d = −0.68), A–E (p = 0.009, Cohen’s d = −0.68), B–A (p < 0.001, Cohen’s d = −2.00), B–E (p < 0.001, Cohen’s d = −0.88), C–A (p = 0.011, Cohen’s d = −0.67), E–A (p = 0.001, Cohen’s d = −0.79), and E–B (p < 0.001, Cohen’s d = −1.06).
Dataset 1—left M1
TMS applied to the left M1 produced significant changes in the ROF for four of the five microstates within 500 ms post-pulse (figure 2(e)). Microstate A showed a significant increase following M1 stimulation (Cohen’s d = 0.77), contrasting with its decrease observed after DLPFC stimulation. Microstate D also exhibited an increase in ROF, albeit with a smaller effect size (Cohen’s d = 0.10). Conversely, Microstates B and E displayed significant decreases (Cohen’s d = −0.59 and −0.76, respectively), while Microstate C showed no significant change.
The analysis of RTF during the 20–500 ms interval following left M1 stimulation revealed a specific pattern of alterations in microstate transitions (figure 2(f)). Four microstate transitions showed significant changes. Transitions between microstates B and E were bidirectionally reduced: B-E (p < 0.001, Cohen’s d = −0.87) and E–B (p < 0.001, Cohen’s d = −0.82). In contrast, transitions between microstates A and D were significantly enhanced: A–D (p < 0.001, Cohen’s d = 1.06) and D–A (p = 0.006, Cohen’s d = 0.68). Comprehensive test statistics for these analyses are provided in table S1.
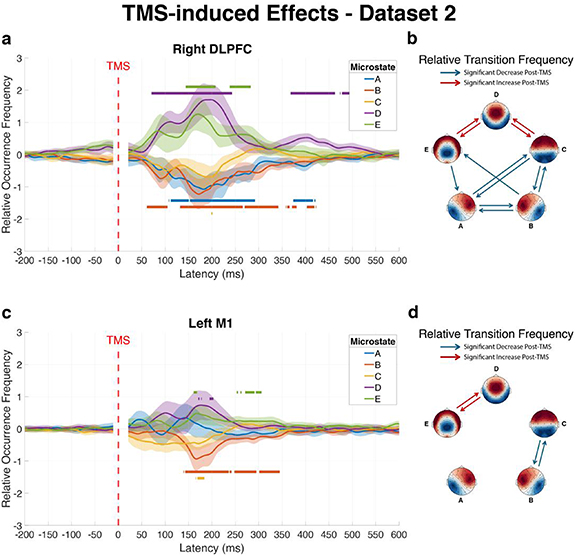
3.3. Testing the generalizability of results using an independent datasetIn dataset 2, right DLPFC stimulation produced significant alterations in microstate dynamics that largely replicated our primary findings with dataset 1. Analysis of ROF revealed significant decreases in microstates A (Cohen’s d = −1.08) and B (Cohen’s d = −1.02) alongside significant increases in microstates D (Cohen’s d = 1.18) and E (Cohen’s d = 1.06) within the first 500 ms post-pulse (figure 3(a)). Microstate C showed no significant changes in ROF in this independent sample.
Figure 3. Replication of significant alterations in microstate dynamics following TMS in an independent cohort (dataset 2). The figure confirms the reproducibility of changes in brain microstates following TMS using an independent dataset, focusing on right DLPFC (a) and (b) and left M1 (c) and (d) stimulation sites. Left panels (a) and (c) show the average ROF trajectories of each microstate over time across subjects, with shaded areas representing 95% confidence intervals. Horizontal lines above or below curves indicate time intervals with statistically significant increases or decreases relative to baseline, respectively. Right panels (b) and (d) display matrices of significant changes in RTF between microstate pairs during the post-pulse window (20–500 ms) compared to baseline, where red connections indicate significantly increased transitions and blue connections represent significantly decreased transitions.
Download figure:
Standard image High-resolution imageFor the RTF following right DLPFC stimulation in dataset 2, eight transition types showed significant decreases in the early post-pulse window (+20 to +500 ms relative to pulse onset, figure 3(b)): A–B (p < 0.001, Cohen’s d = −1.79), A–C (p = 0.001, Cohen’s d = −1.12), B–A (p < 0.001, Cohen’s d = −1.77), B-C (p < 0.001, Cohen’s d = −1.30), B-E (p = 0.044, Cohen’s d = −0.79), C–A (p = 0.001, Cohen’s d = −1.16), C–B (p < 0.001, Cohen’s d = −1.31), and E–A (p = 0.009, Cohen’s d = −0.94). Conversely, four transition types showed significant increases: C–D (p = 0.020, Cohen’s d = 0.87), D–C (p = 0.007, Cohen’s d = 0.97), D–E (p < 0.001, Cohen’s d = 1.66), and E–D (p < 0.001, Cohen’s d = 1.57).
In dataset 2, stimulation of the left M1 resulted in ROF changes, including significant decreases in microstates B (Cohen’s d = −1.24) and C (Cohen’s d = −1.05), and significant increases in microstates D (Cohen’s d = 0.80) and E (Cohen’s d = 0.91) within 500 ms post-pulse (figure 3(c)). Microstate A showed no significant ROF changes in this independent sample.
The transition analysis for left M1 stimulation revealed (figure 3(d)) significant changes only in the early post-pulse window, including transitions B–C (p < 0.001, Cohen’s d = −1.89) and C–B (p < 0.001, Cohen’s d = −1.28) showing significant decreases, while transitions D–E (p = 0.005, Cohen’s d = 1.00) and E–D (p = 0.016, Cohen’s d = 0.89) exhibited significant increases. Comprehensive statistical details for both stimulation conditions are provided in table S3.
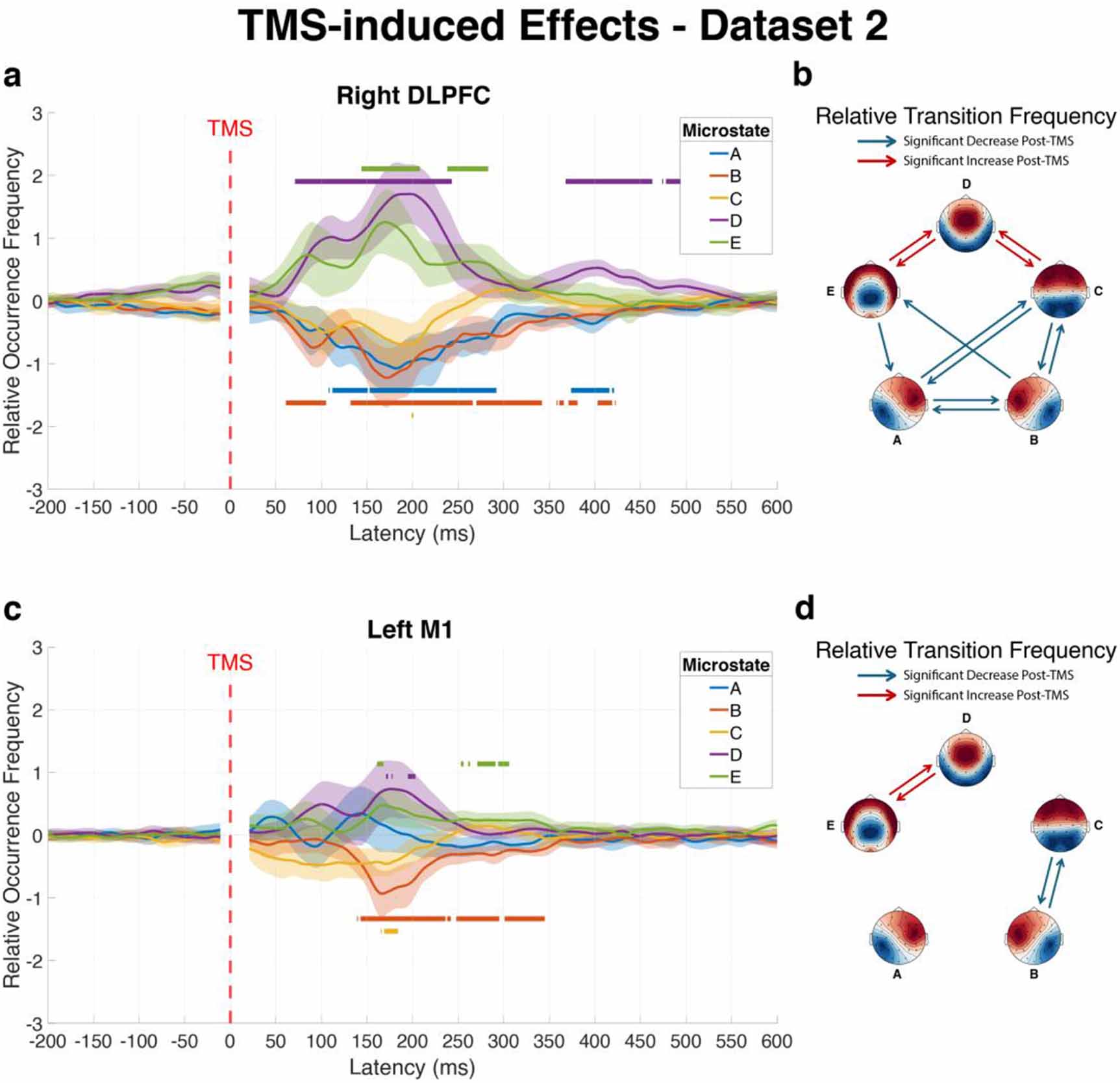
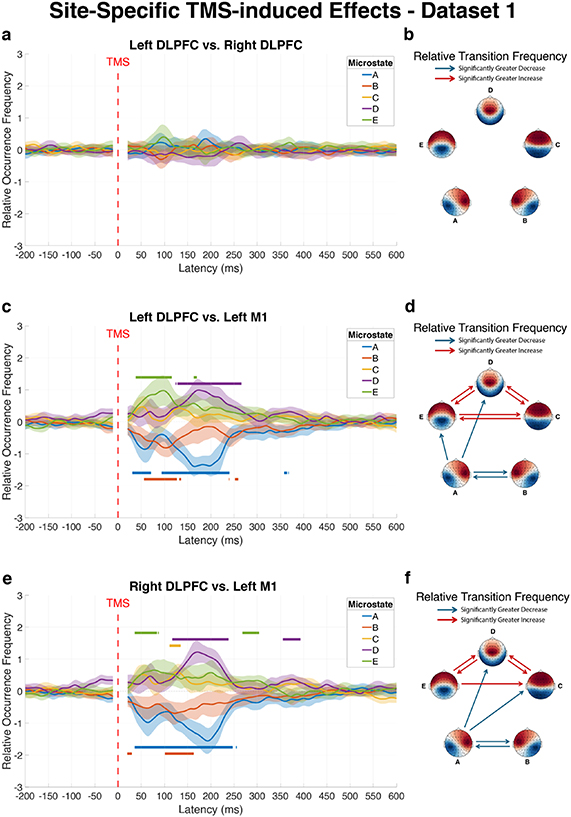
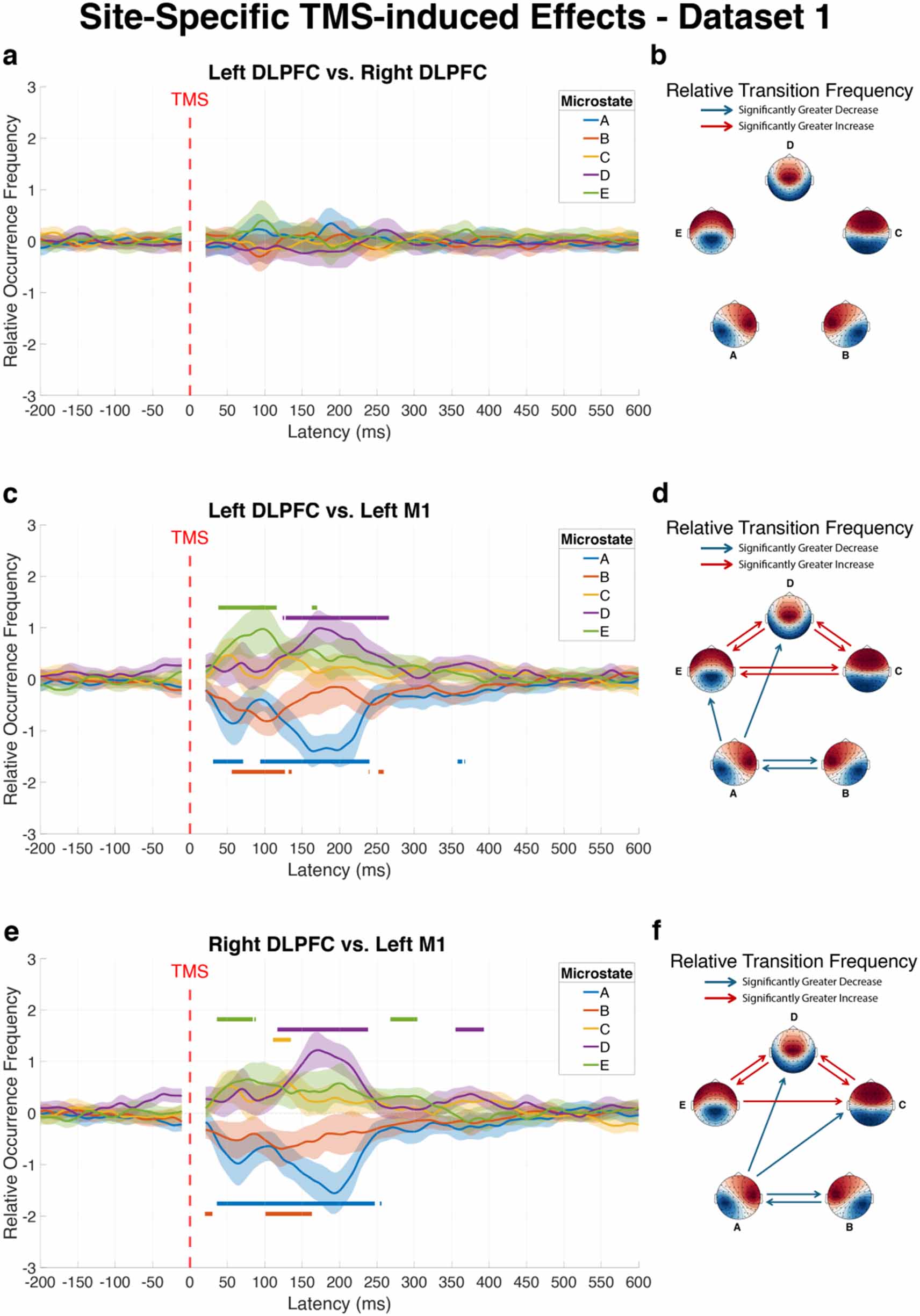
3.4. Site-specific effects of TMS on EEG microstate dynamicsTo determine whether different cortical targets produce distinct effects on global brain network dynamics, we conducted direct comparisons between stimulation sites using dataset 1, allowing us to identify whether stimulation of different cortical regions produced distinct patterns of microstate dynamics.
Left DLPFC versus right DLPFC
Comparison between left and right DLPFC stimulation revealed no significant differences for changes in microstate dynamics following TMS. Neither the ROF of individual microstates nor the RTF between microstate pairs differed significantly between these two stimulation sites (figures 4(a) and (b)). Given that no hemisphere-specific effects were observed for DLPFC stimulation, results from left and right DLPFC are interpreted collectively as DLPFC effects in the discussion.
Figure 4. Differential effects of stimulation site on microstate dynamics following TMS. The figure illustrates direct comparisons of microstate modulations between different cortical targets in dataset 1: left DLPFC vs right DLPFC (a) and (b), left DLPFC vs left M1 (c) and (d), and right DLPFC vs left M1 (e) and (f). Left panels (a), (c) and (e) show differences in average ROF trajectories of each microstate over time across subjects, with shaded areas representing 95% confidence intervals. Horizontal lines above curves indicate time intervals where the first stimulation site produced significantly greater microstate occurrence than the second site, while lines below curves indicate intervals where the first site induced significantly lower occurrence compared to the second site, as determined by TFCE analysis. Right panels (b), (d) and (f) display matrices highlighting significant differences in RTF between microstate pairs during the post-pulse window (20–500 ms), where red connections indicate significantly greater transition probabilities for the first stimulation site compared to the second, while blue connections represent significantly reduced transition probabilities for the first site relative to the second, all after Bonferroni correction.
Download figure:
Standard image High-resolution imageLeft DLPFC versus left M1
Stimulation of the left DLPFC versus the left M1 produced markedly different effects on microstate dynamics (figure 4(c)). Within the first 500 ms post-pulse, microstates A and B showed significantly lower occurrence following left DLPFC stimulation compared to left M1 stimulation (Cohen’s d = −1.50 and −1.13, respectively). Conversely, microstates D and E occurred significantly more frequently after left DLPFC stimulation (Cohen’s d = 1.35 and 0.92, respectively).
The transition analysis revealed substantial differences between these stimulation sites (figure 4(d)). Ten microstate transitions showed significant differences in the early window (20–500 ms). Transitions involving microstates A and B showed greater reductions after left DLPFC stimulation: A–B (p < 0.001, Cohen’s d = −0.96), A–D (p = 0.004, Cohen’s d = −0.72), A–E (p = 0.006, Cohen’s d = −0.69), and B–A (p < 0.001, Cohen’s d = −1.02). In contrast, transitions involving microstates C, D, and E showed greater increases following left DLPFC stimulation: C–D (p < 0.001, Cohen’s d = 1.02), C–E (p = 0.015, Cohen’s d = 0.64), D-C (p < 0.001, Cohen’s d = 0.97), D–E (p < 0.001, Cohen’s d = 1.06), E–C (p = 0.024, Cohen’s d = 0.61), and E–D (p < 0.001, Cohen’s d = 0.86).
Right DLPFC versus left M1
Comparison between right DLPFC and left M1 stimulation produced a pattern similar to the left DLPFC and M1 comparison (figure 4(e)). Microstates A and B occurred significantly less frequently following right DLPFC stimulation compared to left M1 stimulation (Cohen’s d = −1.55 and −0.91, respectively), while microstates D and E showed significantly higher occurrence following right DLPFC stimulation relative to left M1 stimulation (Cohen’s d = 1.88 and 1.13, respectively).
The transition analysis (figure 4(f)) revealed nine microstate transitions with significant differences. Four transitions showed greater decreases after right DLPFC stimulation: A–B (p = 0.003, Cohen’s d = −0.77), A–C (p = 0.007, Cohen’s d = −0.73), A–D (p = 0.001, Cohen’s d = −0.83), and B–A (p = 0.002, Cohen’s d = −0.82). Five transitions showed greater increases after right DLPFC stimulation: C–D (p < 0.001, Cohen’s d = 1.08), D–C (p < 0.001, Cohen’s d = 1.00), D–E (p < 0.001, Cohen’s d = 1.08), E–C (p = 0.031, Cohen’s d = 0.63), and E–D (p < 0.001, Cohen’s d = 0.86). Full statistical details are available in table S4.
The relationship between local cortical activity and ongoing global brain state dynamics remains poorly understood, particularly how localized perturbations can rapidly propagate to influence large-scale networks. Our study demonstrates that TMS-induced local perturbations affect EEG microstates in a distinctly region-specific manner, supporting our hypothesis that stimulation of regions belonging to different large-scale functional networks selectively modulates their associated global brain states within milliseconds. Specifically, stimulation of the left and right DLPFC sites, analyzed separately, was associated with increased occurrence and transitions of microstate D, consistent with its role in the frontoparietal network and executive-related processing, while M1 stimulation enhanced Microstate A, which has been linked to sensory-motor circuits [44]. These effects were robust: microstate changes induced by both DLPFC and M1 stimulation replicated across repeated sessions (detailed in supplementary materials), while the DLPFC finding and region specificity results were further validated in an independent dataset. Together, these findings demonstrate a double dissociation with respect to how stimulation of two distinct cortical regions led to differing (and in some cases, opposing) changes to EEG microstate dynamics. Our findings thereby provide novel insight into how localized perturbations to the ongoing activity of a single brain region can rapidly propagate and alter distinct global brain states. This causal understanding of region-to-global brain state communication provides a foundation for neuromodulation approaches to target specific aspects of large-scale brain dynamics and organization.
Prior TMS-EEG studies have characterized stimulation effects using several complementary approaches. Prior analyses of temporal and frequency components of trial-averaged TEPs, across sensors or source estimates, have focused on quantifying changes in the strength of cortical reactivity in the time or frequency domain [26, 65]. These studies have reported distinct TEP topographies across different brain regions, suggesting that time-frequency-based analysis of TEPs may not fully capture their underlying sources [26]. Connectivity-based methods extend inference to trial-level inter-regional interactions: directed transfer function analyses estimate frequency-specific directional information flow, and phase-locking value measures quantify changes in inter-regional phase synchronization [27, 31]. Although informative about coupling, these approaches represent dynamics as sets of pairwise relationships rather than integrated global configurations, making it difficult to determine whether stimulation biases the brain toward particular whole-brain states or alters switching between them. Recently, microstate analysis was applied in a TMS-EEG study following stimulation of a single cortical site. Performed on trial-averaged TEPs, this study addressed how stimulation parameters shape the evoked microstate sequence [40]. When derived from averaged responses, microstates remain dominated by phase-locked activity and do not capture trial-specific induced dynamics that may reflect reconfiguration of ongoing state activity. In contrast, we apply microstate analysis at the single-trial level, preserve trial-to-trial variability, and quantify changes in frequencies of microstate occurrence and transitions relative to the pre-pulse baseline, providing a direct test of how stimulation modulates ongoing global brain-state dynamics. Finally, by stimulating three cortical sites spanning two functionally distinct regions (left and right DLPFC and left M1), we show that these effects are site-specific, demonstrating that focal perturbations can differentially bias the brain’s moment-to-moment global state sequence within milliseconds.
The ability of local perturbations by TMS to rapidly affect ongoing global brain states likely arises from the propagation of activity from the targeted region. The propagation of activity to distant regions following TMS to a single brain region is well established across multiple modalities, including EEG [19, 25, 65], fMRI [66], and intracranial recordings [67, 68]. This evoked activity is thought to involve transsynaptic effects at the site of stimulation that can either excite or inhibit local intracortical circuits, which then propagate and influence distal regions [69]. In this context, our findings show that localized cortical stimulation can induce rapid changes in global brain state dynamics, as evidenced by changes in EEG microstate features following stimulation of different cortical regions. Furthermore, the onset of microstate changes broadly aligns with reported cortical reactivity responses and their propagation as measured by TEPs [70]. Studies suggest the initial cortical response to be strongest at or near the stimulated region (approximately up to 50 ms following TMS), then followed by the emergence of a global response, believed to reflect recurrent and orchestrated activity from regions and networks distal to the stimulated area [71, 72]. These microstate changes were also present for the first 500 ms, but not after, which also aligns with the commonly reported timing of when TMS-evoked cortical reactivity returns to baseline values. Thus, our results go beyond conventional evoked response measures, offering novel evidence that local perturbations can fundamentally alter the organization of large-scale brain activity.
Following stimulation of the left and right DLPFC, the largest changes to EEG microstate dynamics were a significant increase in the occurrence and transitions of microstate D and a decrease in the occurrence and transitions of microstates A and B. TMS applied to a node within a specific functional network has been found to elicit significantly greater responses within the stimulated network compared to non-stimulated networks [25], suggesting that stimulating a region may enhance global brain states or networks that the targeted region belongs to. As the DLPFC is a critical node of the frontoparietal network, the observed changes in microstate dynamics may reflect increased frontoparietal network engagement. Although EEG microstates and canonical functional networks are not directly equivalent, they show empirical associations [36, 43, 44, 73, 74]. This includes correlations between Microstate D and the frontoparietal network [43, 44, 73]. Consistent with this netw
Comments (0)