
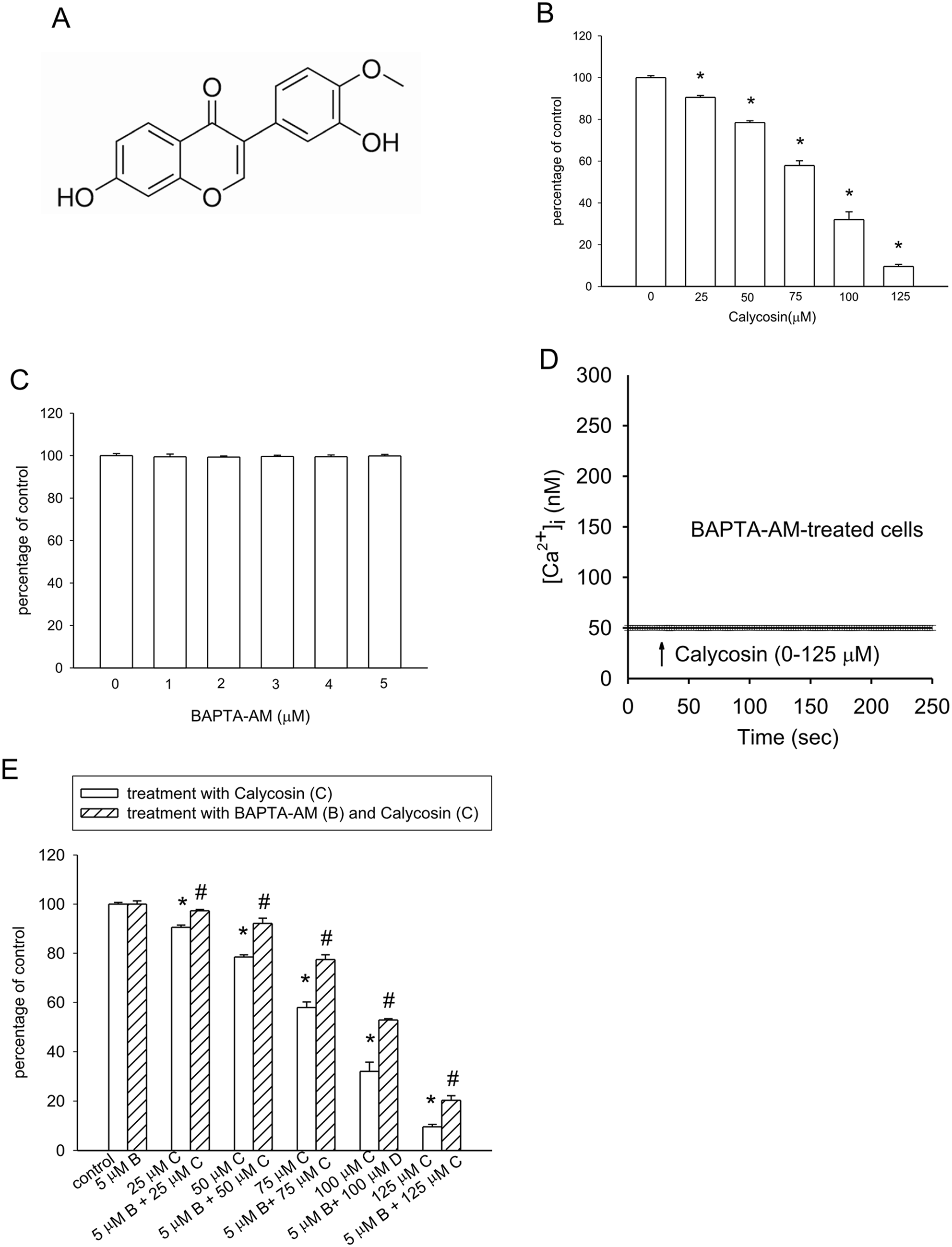
Remember me

Breast cancer represents one fourth of all cancer cases in women worldwide [37], underscoring the urgent need for more effective and durable therapies. While immune checkpoint inhibitors (ICIs) targeting CTLA 4 and PD 1 have revolutionized cancer treatment, their application in breast cancer has shown limited success, partly due to tumor-intrinsic resistance mechanisms. CD200 is one such inhibitory ligand that is frequently overexpressed on breast tumors, where it binds CD200R on antigen presenting cells (APCs) and lymphocytes to drive immune evasion [25,26,27]. Unlike CTLA-4, absence of CD200 signaling does not induce autoimmunity in knockout mice, suggesting a potentially safer checkpoint target [29].
Studies across various cancer models have shown that CD200/CD200R signaling can be therapeutically targeted using monoclonal antibodies (mAbs) [35]. The humanized anti-CD200 mAb, known as Samalizumab, has completed early-phase of clinical trial in hematological malignancies [24]. Similarly, a first-in-human study of 23ME-00610, an anti-CD200R1 antibody, demonstrated an acceptable safety and favourable pharmacokinetics in patients with advanced solid malignancies [38]. Preliminary clinical activity was also observed, further supporting the therapeutic potential of CD200/CD200R blockade [38].
We previously reported that local HT delivered via near-infrared radiation (NIR) delayed tumor progression, induced tumor necrosis, and increased infiltration of CD8+ T cells [34]. Mechanistically, HT induces heat shock protein 70 (Hsp70), which enhances tumor antigen uptake and dendritic cell (DC) maturation. Additionally, HT transiently improves tumor perfusion and vascular permeability, facilitating immune cell extravasation [39]. It also promotes the releasing damage-associated molecular patterns (DAMPs) like Hsp70, which activate APCs and stimulate pro-inflammatory cytokines to create a chemotactic gradient for effector lymphocyte trafficking [40].
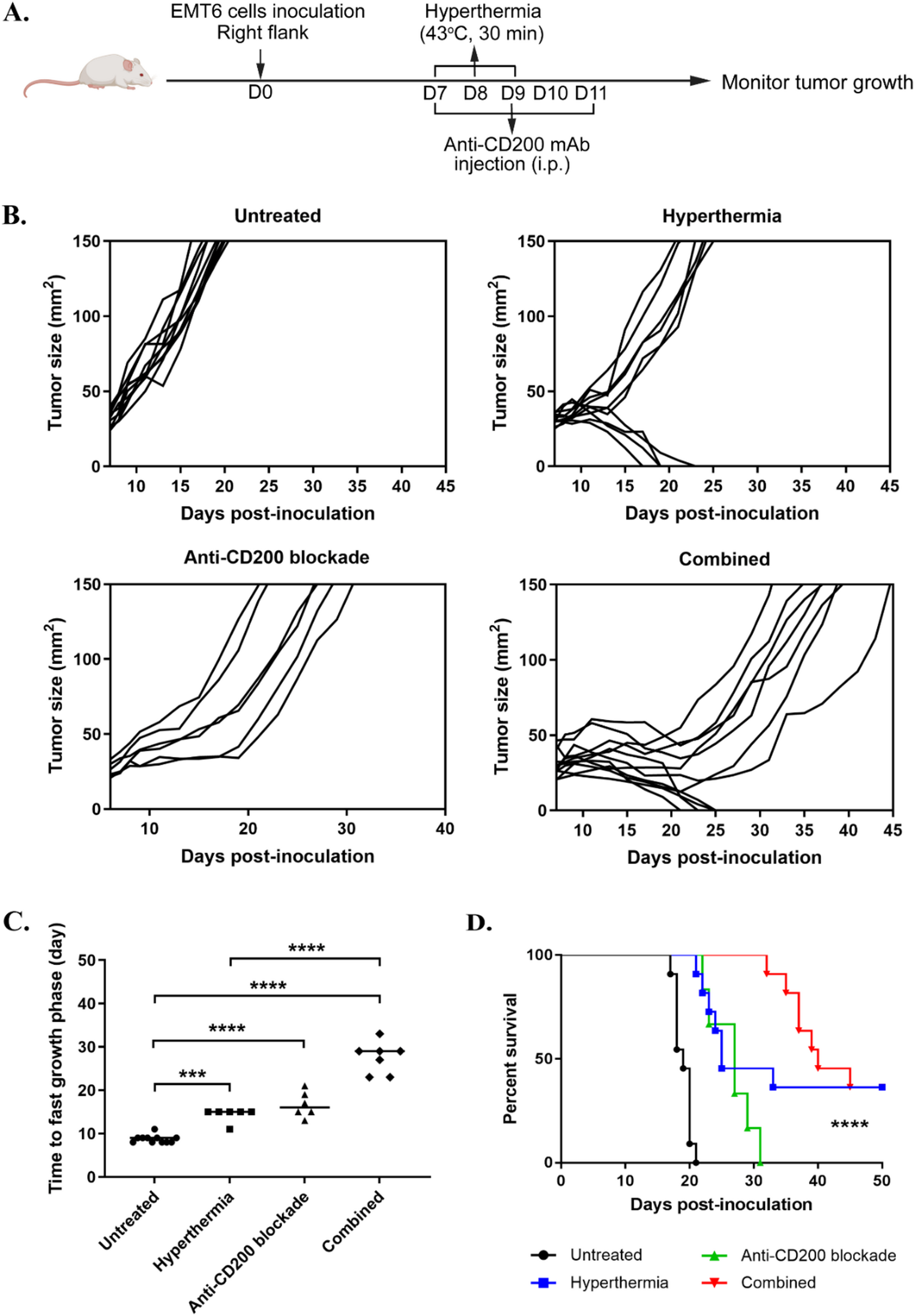
In this study, we present the first preclinical evaluation of combined local HT and anti-CD200 mAb treatment in the EMT6 breast cancer mouse model. We observed that the addition of anti-CD200 mAb to HT (1) further delayed tumor progression and extended median survival compared to monotherapy; (2) enhanced activation and infiltration of CD8⁺ T cells and NK cells; and (3) reduced immunosuppressive Tregs and MDSCs in the tumor microenvironment (TME). These findings suggest that HT primes an immune-permissive TME that enhances the effectiveness of CD200 blockade. Figure 6 summarizes the hypothesized mechanisms underlying these observations, which are proposed based on established literature and supported by phenotypic and cytokine-based immune profiling rather than direct functional assays.
In breast cancer, immune checkpoint blockade has shown limited efficacy as monotherapy, emphasizing the need for combination strategies that modify the tumor microenvironment. Consistent with existing literature, hyperthermia acts as an immune-priming modality by enhancing antigen presentation and immune cell infiltration, and our data show that combining hyperthermia with CD200 checkpoint blockade further augments effector immune responses while reducing immunosuppressive cell populations. These findings support the rationale that immune-priming approaches such as hyperthermia can enhance the effectiveness of checkpoint inhibition in breast cancer.
Anti-CD200 therapy is thought to disrupt inhibitory signaling on CD200R-expressing APCs and lymphocytes, thereby restoring effector function and enhancing DC-T cell priming (Fig. 6A). In an HT-primed TME, this blockade may further amplify cytotoxic T lymphocyte (CTL) activity and limits the expansion of Tregs. This is supported by our observation of increased CD45⁺ lymphocyte density and CD8⁺ T cell infiltration (Fig. 6B), along with decreased Treg infiltration (Fig. 6C), suggesting enhanced CD8+ T cells effector function and a shift toward an effector phenotype (Fig. 6D). In addition, blocking CD200 may prevent suppression of NK and T cells by CD200-expressing tumor on immune cells, thus preserving their cytotoxic function (Fig. 6E).
Given that T cells also express CD200, blocking CD200/CD200R signaling is important to ensure that increased in TIL expressing CD200 does not shifted T cells activity towards immunosuppressive phenotype. Engagement of CD200 on T cells with CD200R on APC expressing CD200R could induce tolerogenic DCs, which subsequently promote Tregs development [9]. This is supported by Li et al. (2012), who reported that CD200/CD200R signaling influences T cells differentiation, and increased signaling through CD200R1 may bias toward Tregs [41]. Moreover, unchecked CD200 signaling on lymphocytes may lead to T cell overactivation, chronic inflammation and MDSC expansion. Our results highlight that blocking this signaling helps re-establish immune homeostasis, especially when coupled with HT treatment.
Fig. 6
Proposed illustration summarizing the effect of local HT treatment combined with anti-CD200 blockade therapy on TME
In addition to modulating T cells, the combination therapy significantly increased intratumoral NK cells (Fig. 6F). This enhancement can be attributed to a dual mechanism. Firstly, HT-induced expression of Hsp70 facilitates NK cell recognition and cytotoxicity, as Hsp70 presents surface epitopes that are directly recognized by NK cells [42]. However, current study shows markedly increased of NK cells infiltration when CD200 was simultaneously blocked. This is because CD200 has an inhibitory role on NK cells by suppressing their cytotoxic function and reducing IFN-γ production [20]. Therefore, blocking CD200 prevents this suppression, allowing NK cells to retain their function sustain IFN-γ secretion [21]. This dual activation mechanism highlights the potential of combining HT with CD200 blockade to boost both innate and adaptive immune responses against tumors.
Increased cytolytic activity of CTLs and NK cells is essential for mounting an effective anti-tumor response. However, suppressing immunosuppressive cells, particularly MDSCs, is equally critical, as they are key contributors to ICI resistance and immune escape [43]. While it is established that CD200/CD200R engagement inhibit general myeloid cell activity, paradoxically, this effect is stimulatory rather than suppressive for MDSC [12]. Upon CD200R engagement, MDSCs secrete granulocyte colony-stimulating factor (G-CSF) and granulocyte-macrophage colony-stimulating factor (GM-CSF), which further enhance their suppressive function and promote tumor angiogenesis, progression, and metastasis [44, 45].
Blocking CD200/CD200R signaling is thus hypothesized to limit MDSC expansion in which a study by Choueiry et al. demonstrated CD200 blockade reduced MDSC levels in pancreatic tumor [46]. Consistent with this, our study showed a significant reduction in MDSCs with anti-CD200 treatment, particularly when combined with HT (Fig. 6G). In contrast, MDSC levels in control and HT groups were similar, indicating that HT alone is insufficient to suppress MDSC activity. Combining HT with CD200 blockade may thus offer a therapeutic advantage, especially when considering that HT alone, while effective in enhancing immune cell infiltration and cytotoxic activity, does not reliably suppress MDSC expansion. Tumors exploit MDSCs to maintain an immunosuppressive environment, enabling resistance to immune attack even after treatments that boost effector cell activation and function [47]. Therefore, this combination strategy addresses both effector activation and immune evasion, promoting a more effective anti-tumor response.
While our results demonstrate promising immunological shifts and therapeutic benefit, this preliminary evaluation has limitations. Despite having measured key intratumoral cytokines (IL 2, IFN γ, TNF α, IL 10) and chemokine CXCL2, the observed differences were mostly non-significant. Additionally, functional assays such as cytotoxicity or signaling analyses were not performed, limiting direct confirmation of effector cell function. Future studies should expand cytokine panels and assess temporal dynamics at multiple time points to fully validate the proposed mechanism. Additionally, formal synergy analyses, such as combination index calculations, could quantify interactive effects beyond our additive observations. Most importantly, this work represents a single exploratory experiment in the EMT6 model, and thus should be interpreted as a pilot study. Replication in larger cohorts and different models is essential to validate these effects.
Given that CD200 is broadly expressed on various leukocyte subsets including T cells, B cells, NK cells, and DCs, systemic administration of anti-CD200 mAb may impact immune regulation beyond the tumor site. This raises safety considerations: altering CD200 signaling in the periphery may shift activation thresholds, potentially inducing off-target inflammatory effects. Future studies should therefore include comprehensive profiling of peripheral blood and secondary lymphoid organs, as well as monitoring for signs of immune-related adverse events, to fully evaluate the safety and systemic impact of this combination therapy.
Comments (0)