Reduced JUP expression correlates with advanced nodal disease and poor survival
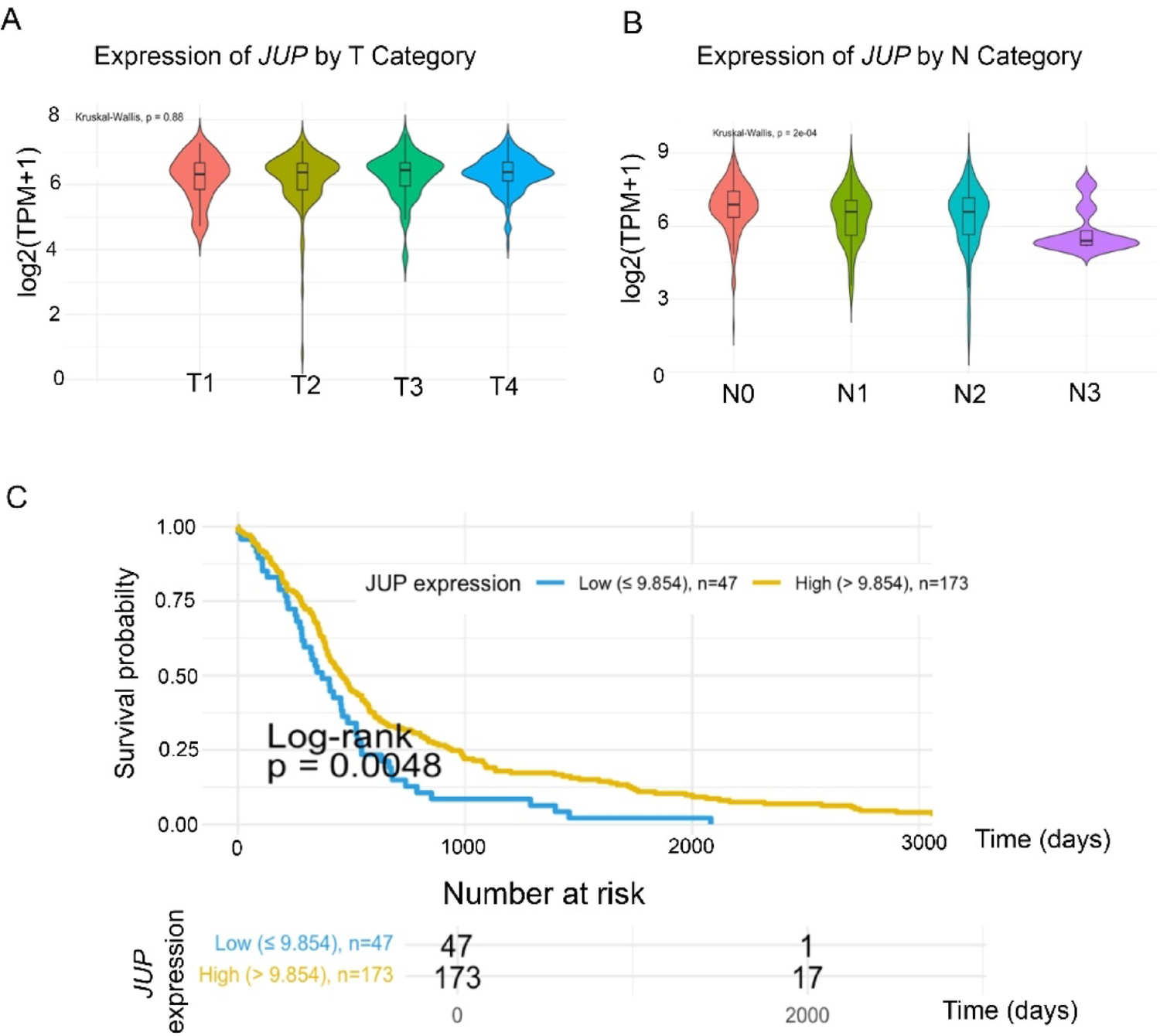
To assess the clinical relevance of JUP expression in head and neck squamous cell carcinoma (HNSCC), we analyzed RNA-sequencing data from 564 tumor and normal tissue samples in the TCGA-HNSCC cohort.
While no statistically significant differences in JUP expression were observed across tumor T stages (Fig. 1A; p = 0.88). JUP expression (log₂[TPM + 1]) was significantly associated with lymph node involvement (p = 0.0002; Fig. 1B). Mean JUP expression was highest in N0 tumors (10.522 ± 0.693) and progressively decreased in more advanced nodal stages, with the lowest levels detected in N3 tumors (5.816 ± 1.143). These data suggest that JUP expression is inversely correlated with nodal metastasis and may be involved in the regulation of lymphatic dissemination.
We next assessed whether JUP expression levels are associated with overall survival (OS). Kaplan-Meier analysis was performed for 220 primary HNSCC tumors with available survival data. Optimal cut-off values were determined using the maximally selected rank-statistics method, controlling for multiple testing. The identified JUP expression threshold was 9.854 (log₂[TPM + 1]); patients with lower expression exhibited significantly reduced OS (p = 0.0048; Fig. 1C). These results support a prognostic role for JUP in HNSCC.
JUP knockdown enhances epithelial wound closure
To explore the functional significance of JUP in tumor cell behavior, we performed siRNA-mediated knockdown in FaDu cells. Efficient silencing of JUP was confirmed by Western blot analysis (Fig. 2A, B; control: 3.05 ± 0.42% vs. JUP siRNA: 1.31 ± 0.42%, p < 0.01). In scratch wound assays, JUP-depleted FaDu cells exhibited a markedly accelerated wound closure after 24 h and 48 h compared with both control and non-targeting (nt) control cells (Fig. 2C, D). After 24 h, wound closure reached 44.9 ± 7.3% in JUP-knockdown cells, compared with 24.6 ± 6.8% in controls and 27.8 ± 5.7% in nt controls (p < 0.05). At 48 h, wound closure was 45.1 ± 6.2% in JUP-siRNA cells, compared with 32.8 ± 7.6% in controls and 33.0 ± 5.9% in nt controls (p < 0.01), confirming a sustained motility-enhancing effect following JUP depletion.
To further validate these findings in an additional cell model, we performed complementary experiments in HPV-positive UPCI-SCC154 cells. After establishing an efficient JUP knockdown using siRNA (Fig. 2E, F), we next conducted scratch wound assays to assess the effect of JUP depletion on cell motility. Consistent with our previous results in FaDu cells, JUP-silenced SCC-154 cells exhibited markedly accelerated wound closure. After 24 h, wound closure was 19.7 ± 4.6% in JUP-siRNA cells, 12.5 ± 5.2% in nt controls, and 13.8 ± 4.3% in controls (p < 0.05). After 48 h, wound closure reached 44.9 ± 7.3% in JUP-knockdown cells, compared with 24.6 ± 6.8% in controls and 27.8 ± 5.7% in nt controls (p < 0.01). Collectively, our findings demonstrate that JUP depletion induces consistent phenotypic alterations across both HPV-negative (FaDu) and HPV-positive (SCC154) HNSCC cell models, suggesting a broader functional role of JUP in regulating tumor cell properties independent of HPV status.
Increased Ki-67 expression indicates enhanced proliferation following JUP knockdown
To determine whether the increased wound closure observed upon JUP loss was associated with enhanced cell proliferation, we examined Ki-67 expression as a proliferation marker.
Immunostaining 24 h after scratch, revealed a significantly higher percentage of Ki-67–positive cells in JUP-silenced monolayers (2.67 ± 0.78%) compared with controls (1.33 ± 0.46%; p < 0.01; Fig. 3A, B). This finding was further corroborated by Western blot and qPCR analyses, which confirmed increased Ki-67 expression at both the protein- (Fig. 3C, D) and mRNA levels (Suppl. Figure A) following JUP knockdown (mRNA: control = 1.22 ± 1.56 vs. JUP siRNA = 2.87 ± 1.31; p < 0.01). It should be noted that epithelial regeneration and tumor progression generally involve a combination of cell proliferation and motility-related processes [20]. To further dissect the potential contribution of cell migration, we analyzed phosphorylation of focal adhesion kinase (FAK) as a surrogate marker of migratory activity.
Western blot analyses demonstrated significantly elevated FAK phosphorylation in JUP-depleted cells relative to controls values (Fig. 3E).
Collectively, these results indicate that loss of JUP activates pathways linked to both proliferation and migration.
To further explore the mechanisms underlying the accelerated wound closure observed in HPV-positive SCC154 cells, we examined potential changes in cell proliferation.
Immunostaining revealed a significant increase in Ki-67-positive cells following JUP depletion (31.04 ± 4.68%) compared with control (18.78 ± 4.16%) and non-targeting (nt) controls (16.83 ± 3.13%) (p < 0.01; Fig. 3F, G). This observation was further confirmed at the protein level by Western blot analysis, which revealed elevated KI-67 expression, followed by increased FAK phosphorylation after JUP depletion (2.14 ± 3.89) compared with control cells (0.87 ± 3.72; Fig. 3H-J).
Loss of JUP activates PI3K/AKT signaling in HPV-negative and HPV-positive HNSCC cell models
To further investigate the molecular mechanisms by which JUP regulates cellular proliferation and migration, we examined activation of the PI3K/AKT signaling pathway.
Western blot analysis demonstrated that JUP knockdown resulted in increased levels of phosphorylated AKT (p-AKT) and the proliferation marker Ki-67 in both FaDu and SCC-154 cells (Fig. 4A, B).
Quantitative analysis revealed that p-AKT levels increased from 1.40 ± 0.49 in control cells to 2.55 ± 0.94 in JUP-siRNA–treated FaDu cells (Fig. 4C; p < 0.001) and from 1.23 ± 0.74 to 2.31 ± 1.12 in SCC-154 cells (Fig. 4D; p < 0.001).
Similarly, Ki-67 expression was elevated following JUP depletion, increasing from 1.17 ± 0.34 to 2.95 ± 0.83 in FaDu cells (Fig. 4E) and from 0.98 ± 0.57 to 2.68 ± 1.24 in SCC-154 cells (Fig. 4F).
To determine whether these effects were mediated by PI3K/AKT signaling, JUP-depleted cells were treated with the PI3K inhibitor LY294002. In both cell lines, PI3K inhibition markedly reduced p-AKT and Ki-67 levels, restoring them to near-control values (Fig. 4A–F), indicating PI3K/AKT-dependent regulation.
To assess whether these molecular changes translated into functional consequences, scratch wound assays were performed in FaDu cells, which displayed the strongest JUP-dependent activation of PI3K/AKT signaling.
JUP knockdown significantly accelerated wound closure compared with control conditions (Fig. 4G, H). Importantly, treatment with PI3K inhibitor (LY294002) completely abrogated this effect, restoring wound healing rates to control levels.
Together, these findings demonstrate that loss of JUP enhances proliferative and migratory behavior via activation of the PI3K/AKT pathway.


Comments (0)