
Remember me
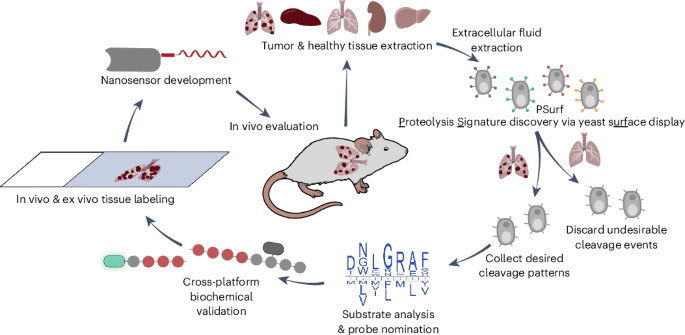
PSurf employs a double-tagged peptide display approach where each peptide displayed on the yeast surface is flanked by two epitope tags recognized by fluorescently labeled antibodies (Fig. 1b). When proteolysis occurs, one tag is cleaved off, allowing tracking of cleavage events by measuring fluorescence change through cell sorting. Yeast populations displaying peptides with specific proteolytic activities are sorted for next-generation sequencing (NGS) analysis or cultured for further rounds of selection.
To optimize the tagging strategy, we used the tobacco etch virus (TEV) protease substrate ENLYFQG. The substrate was fused to the C terminus of Aga2p for yeast surface display, and three tag pairs were compared: HA/cMyc, HA/V5 and V5/HA (N-terminal/C-terminal orientations) (Fig. 1c). Yeast cells displaying the peptides were incubated with TEV protease and analyzed using flow cytometry. Among the designs, the V5/HA tag combination exhibited homogeneous display and dose-dependent and time-dependent reduction in HA signal upon proteolysis (Fig. 1c,d) and was selected as the optimal design for library construction.
To assess whether the display strategy can capture protease substrate preferences, we compared the canonical TEV protease substrate (ENLYFQG) with a non-canonical substrate (ENLYFQV), observing notably reduced cleavage of the latter, as expected41 (Fig. 1d). By contrast, cleavage was similar between substrates when using a TEV protease mutant (TEV L56V, S135G, S219V), which is known to have broader substrate tolerance42 (Fig. 1e).
We next used NNK degenerate codons (N: A/T/C/G, K: G/T; Extended Data Fig. 1a,b) to construct the PSurf library displaying randomized seven-amino-acid peptides, allowing exploration of a large sequence space, including sequences not found in native proteins43. The peptide length of seven amino acids was chosen to balance coverage of central recognition motifs and flanking residues while maintaining sufficient sequence diversity within the practical size constraints of a yeast library44. A yeast surface display plasmid was transformed into Saccharomyces cerevisiae, along with the amplified DNA library containing homologous flanking sequences to enable recombination, generating a library of 3 × 108 transformants. As detected by flow cytometry, 58% of the population expressed both epitope tags (Extended Data Fig. 1c). NGS analysis of 5 × 106 transformants showed nucleotide and amino acid frequencies closely aligned with expectations (Fig. 1f, Extended Data Fig. 1d,e and Supplementary Table 1).
To understand the population of yeasts lacking the C-terminal HA tag, we examined this population and identified the two primary contributions as stop codons introduced by NNK usage (Extended Data Fig. 1e) and substrates of yeast proteases (Extended Data Fig. 3a,b). Of this population, 34% encoded full-length peptides with strong enrichment of LxxR and RR motifs (Extended Data Fig. 3b). These motifs match the known cleavage sequences of the yeast protease Kex2, indicating that the peptide cleavage occurred during secretion to the cell surface and demonstrating that PSurf can unbiasedly reveal endogenous protease substrate preferences (Extended Data Fig. 3a)32. The library was further sorted to enrich for the uncleaved peptide population, generating the final PSurf library used for subsequent proteolysis profiling studies (Extended Data Fig. 1d,e, right panel).
Identifying cathepsin substrates and pH responsesWe first applied the library to recombinant CTSB, a prognostic biomarker associated with multiple solid tumors45, which has been widely developed as a protease trigger for payload release in ADCs and as a therapeutic target for inhibitor development46,47. An input library size of 2 × 107 was selected to preserve high sequence coverage while remaining compatible with the experimental throughput imposed by cell sorting speed. The library was initially treated with the enzyme at pH 4.4, a condition expected to support maximal CTSB activity. The selected population was subsequently subjected to more stringent conditions with reduced enzyme concentration and incubation time (Fig. 2a, Extended Data Fig. 2a,b and Supplementary Fig. 1). In alignment with previous reports, NGS analysis revealed gradual enrichment of the leucine–valine–glycine (LVG) motif across successive rounds of selection48,49.
Fig. 2: Identifying cathepsin substrates. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, Selection schematic for CTSB substrate identification. b, Flow cytometry plots and sequence logos from round 3 of PSurf selection with CTSB at pH 4.4. A strongly enriched LVG cleavage motif was observed under stringent selection conditions (10 nM CTSB). Sorting gates are indicated by dashed lines. Sequence logos indicate amino acids abundance at each position of the seven amino acids substrates (P1–P7). c, Flow cytometry plots and sequence logos showing substrate cleavage motifs after round 3 of selection at pH 6.4 and pH 7.4. Unlike the pH 4.4 results, motifs such as RR and VR were strongly enriched. Sorting gates are indicated by dashed lines. d, z-score plots showing dipeptide enrichment profiles for CTSB at different pH levels. The y axis represents the first amino acid; the x axis represents the second amino acid. Distinct dipeptide motifs were enriched under each pH condition. e, On-yeast cleavage activity of CTSB with newly identified substrates. RRLYAFL and RMLVGFV were highly sensitive to CTSB at both pH 4.4 and pH 6.4; LVGVLLF showed significantly selective activity at pH 4.4 and VRFLAAA at pH 6.4. Yeast populations displaying these peptides were incubated with 70 nM CTSB for 72 hours. Cleavage activity is shown as the −log of HA–AF488 signal normalized to peptide expression (V5–AF647 signal) and to untreated controls. Data represent the mean of n = 3 replicates; error bars indicate standard deviation. STSSTSS served as a no-activity control, and GRLVGFD49 served as a positive control. Unpaired two-tailed t-test was used for pH-dependent activity comparisons, ****P < 0.0001 for LVGVLLF and *P = 0.0456 for VRFLAAA. f, Schematic of the probe and cleavage assay comparing RRLYAFL to the conventional substrate KSGGGK. A synthetic peptide labeled with FAM/CPQ2 was used; cleavage releases the fluorophore, generating a fluorescence signal. RRLYAFL showed enhanced cleavage by CTSB. Data represent the mean of n = 3 replicates. RFU, relative fluorescence units.
pH sensitivity is an important characteristic of cathepsins, enabling them to modulate activity across cellular compartments. We next investigated the pH-dependent substrate preferences of CTSB by performing similar selection cycles at pH 5.4, pH 6.4 and pH 7.4 (Fig. 2c and Extended Data Fig. 2c). Compared to the pH 4.4 selections, smaller library population was cleaved at higher pH values, reflecting the expected decrease in CTSB activity50. NGS results indicated that, as pH increased, enrichment of the LVG motif decreased, whereas RR and VR motifs became more prominent (Fig. 2d and Extended Data Fig. 2d,e). RR and VR were previously identified as sensitive human CTSB substrates at pH 7.0 and pH 6.4, in line with our findings51. Notably, a VR-derivative peptide, valine–citrulline (Val–Cit), is one of the most widely used ADC linkers52. Citrulline is a deaminated form of arginine. Val–Cit is preferred over VR in ADCs because it is more stable in human plasma while retaining robust cleavage activity by cathepsins53. These results highlight the ability of PSurf to identify context-specific enzyme cleavage features.
Because cathepsins primarily promote tumor progression in the extracellular space, where pH is markedly above the lysosomal pH but often lower than normal tissue, there is a strong interest in identifying novel cathepsin substrates or peptide-based inhibitors that function at tumor microenvironmental pH to better target tumor-associated cathepsins. We nominated four peptides from the pH 6.4 screen to be displayed individually on yeast and tested their CTSB sensitivity (Fig. 2e and Supplementary Table 2). Consistent with the motif enrichment results, the VR-containing peptide VRFLAAA showed selectivity at pH 6.4, whereas the LVG-containing peptide LVGVLLF showed selectivity at pH 4.4 (Fig. 2e). Peptides containing motifs enriched at both pH 4.4 and pH 6.4 maintained activity across pHs, as seen with peptide RMLVGFV (motif RM) and peptide RRLYAFL (motif RLY) (Fig. 2b,c). From our analysis, the novel substrate RRLYAFL emerged as the most enriched sequence in the pH 6.4 NGS dataset, representing 12% of all sequences. To assess its activity relative to known cathepsin substrates, we designed synthetic fluorogenic substrates, consisting of seven amino acids flanked by a fluorophore and quencher, where cleavage leads to time-dependent fluorescence increase (Fig. 2f, left). Testing this probe alongside a CTSB-responsive probe, the newly identified substrate exhibited faster cleavage kinetics than the known CTSB substrate at an acidic extracellular pH characteristic of the TME (6.4) (Fig. 2f, right). These results demonstrate that our PSurf library can effectively identify both known and novel cathepsin substrates.
Samples derived from living organisms exhibit greater complexity than purified proteases due to the presence of diverse protease families, endogenous inhibitors, competing substrates, metal ions and other components that modulate protease activity. To evaluate the system’s ability to characterize biologically complex samples, we applied PSurf to a mouse upper intestinal washout sample, which resulted in cleavage of 24% of displayed peptides in 24 hours and 10% of peptides in a subsequent 4-hour selection cycle (Extended Data Fig. 3c–e and Supplementary Fig. 2). In agreement with the expected high concentrations of trypsin in this sample, NGS analysis revealed clear arginine enrichment in the cleaved pool and further arginine enrichment across all seven amino acid positions in the more stringent selection. These results demonstrate that PSurf can effectively capture specific proteolytic activities in biological samples and that increased selection stringency enhances enrichment for tissue-sensitive substrates, consistent with observations in the CTSB experiments.
Identification of tumor-selective sequences through differential selectionWe next applied PSurf to tumor samples. We harvested tumor-bearing lungs and healthy organs from two lung metastasis mouse models derived from CT26 cells (colorectal cancer) and B16-F10 cells (melanoma). Both models developed pulmonary metastatic nodules within 10–14 days after intravenous injection. We extracted the soluble fraction of the extracellular matrix (sECM) from healthy tissues (liver, lung, kidney and spleen) and tumor lesions in the lung based on a filter centrifugation method (Fig. 3a and Extended Data Fig. 4a)54. In some contexts, we prepared tissue extracellular samples (ECSs) by gentle mechanical dissociation and collected extracellular proteins55. Both tissue processing methods were carefully performed to minimize altering compartmentalization and regulation or cell lysis, to represent tissue extracellular environments.
Fig. 3: Profiling of tissue extracellular fluids using PSurf. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, Schematic and protocol overview for isolating and profiling tissue-derived extracellular fluids. b, z-score plots showing single amino acid enrichment and depletion across tissues and tumors (liver, lung, kidney, spleen, CT26 CRC lung metastases and B16-F10 lung metastases). Residues with a significant increase compared to pooled mean of z-scores are framed in red, indicating enrichment. Residues with a significant decrease compared to the pooled mean are framed in blue, indicating depletion. Unpaired two-tailed independent t-tests were performed, and P values are provided in Supplementary Table 5. c, Frequencies of enriched amino acids in the input library and each sample type. d, Schematic of the differential selection strategy. The tumor-enriched population was sequentially screened against liver, lung, kidney and spleen samples. After each round, the non-cleaved pool was carried forward to remove substrates cleaved by healthy tissues. The final non-cleaved pool was screened against tumor extracellular fluid to further enrich for tumor-sensitive substrates. e, iceLogos showing per-position amino acid enrichment and depletion before and after differential selection, normalized to the initial library. The pool selected from global profiling was enriched in Leu and Phe, whereas the final pool after differential selection displayed greater sequence diversity. iceLogos were generated using the iceLogo web server. CRC, colorectal cancer. Graphics in a and d created in BioRender; Lord, A. https://biorender.com/5uvic7 (2026).
The PSurf library was incubated with the extracted sECM samples for 24 hours, resulting in substrate cleavage levels of 11% and 10% in CT26 and B16-F10 tumor-bearing lung, respectively, and 7% in healthy lung (Fig. 3a, Extended Data Fig. 4b and Supplementary Fig. 3). Healthy spleen, kidney and liver exhibited cleavage of 8%, 9% and 10%, respectively. The buffer control led to 6% cleavage, reflecting residual precleaved peptides by yeast, although the mean fluorescence intensity (MFI) indicated a reduced degree of cleavage compared to tissue-derived samples. The cleaved fractions were harvested; precleaved peptides were removed by sorting; and the resulting yeast pool was then subjected to a second round of selection with tissue sECM samples and analyzed by NGS.
Interestingly, across both healthy tissue and tumor samples, NGS revealed strong enrichment of sequences containing phenylalanine (Phe), leucine (Leu) and other hydrophobic residues (Fig. 3b,c and Extended Data Fig. 4c,d), characteristic of substrates for several serine proteases56. For example, GZMH and CTSG possess chymotrypsin-like activity and preferentially cleave after large hydrophobic residues such as Phe, as evidenced by their efficient cleavage of the Phe–Leu–Phe synthetic substrate57,58,59. Other secreted serine proteases, such as KLK760, NE and PR3 (ref. 61), also exhibit substrate preferences for Leu or Phe. In addition, some MMPs (MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP12, MMP13 and MMP14) have been reported to prefer leucine at the P1′ position16. Thus, the observed enrichment of Phe/Leu-rich sequences may reflect the integrated activity of proteases acting within the sECM of healthy and tumor tissues. These findings suggest that tissue samples may share common cleavage signatures driven by conserved protease substrate preferences.
To distinguish tumor-specific substrates from those commonly cleaved in both tumor and healthy tissues, we leveraged the capacity of yeast display for repeated positive and negative selection, a strategy used in nanobody and antibody discovery to isolate binders targeting specific epitopes or conformational states of antigens62,63,64 (Fig. 3d). We initiated this differential selection process using the cleaved population obtained from the initial tumor sECM screens (Extended Data Fig. 5a). To deplete substrates also cleaved by healthy sECM, this pool was sequentially incubated with sECM from healthy liver, lung, kidney and spleen, collecting the non-cleaved fraction at each step and culturing for the next selection step. Finally, the remaining non-cleaved pool was exposed to tumor-derived sECM, and the resulting cleaved fraction was collected, enriching for substrates highly sensitive and specific to tumor-associated proteolytic activity (Supplementary Fig. 4).
iceLogos65 generated from cleaved and non-cleaved populations revealed distinct tissue-specific cleavage features (Extended Data Fig. 5b) and clear differences before and after differential selection (Fig. 3e). Although Leu and Phe were highly enriched in the input sequences from both tumor models, these residues were depleted in the output population, where enriched residues included arginine, proline, asparagine, glycine, alanine and aspartate (Extended Data Fig. 6). We selected 10 substrate sequences (Supplementary Table 3) based on several selection criteria: ‘Top hit’ and ‘2nd hit’ are sequences that had the highest NGS reads count for each treatment; ‘Per-position enrichment’ describes sequences comprising the most frequent amino acid in each position; ‘Top unique hit’ is a sequence with the highest NGS reads count not identified in other tissues; and ‘Shared sequence’ was present in all samples. We then tested their tissue specificity on yeast and nominated four peptides preferred by tumor tissue for further biochemical and in vivo analyses (Extended Data Fig. 5c).
Biochemical characterization of PSurf-identified substratesTo further characterize the PSurf-identified sequences, we synthesized these peptides as fluorogenic probes. Lung ECSs from mice bearing CT26 metastases and healthy animals were incubated with the probes to assess differences in substrate cleavage rates (Fig. 4a,b). Lung samples from lipopolysaccharide (LPS)-treated mice were also included to distinguish tumor-specific from inflammation-associated proteolytic activity. Four PSurf-identified substrates were benchmarked against the previously reported MMP substrates PVPLSLVM and PLGLRSW66,67, which contain the conserved MMP cleavage motif PXXL68 as well as AIEFSD, a known substrate of granzyme B69. The kinetic fluorescence assays revealed that all four PSurf-identified probes were cleaved more rapidly in tumor samples than in healthy lung samples and exhibited minimal cleavage in inflamed lung tissue (Fig. 4c). When applied to blood samples from tumor-bearing and healthy mice, we did not detect differences in fluorogenic probe activation (Extended Data Fig. 7a), suggesting that these probes are sensing tissue-level proteolytic changes rather than systemic changes in circulation.
Fig. 4: PSurf-derived substrates show tumor-specific cleavage distinct from canonical MMP substrates. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, Schematic of fluorogenic probes report proteolysis by tissue ECS. b, Kinetic fluorescence traces. DNLGRAF shows higher cleavage in CT26 tumor-bearing versus healthy lung ECS (mean of n = 3). c, Cleavage rates for four PSurf probes, MMP probes (PVPLSLVM and PLGLRSW) and GZMB probe (AIEFDS) in CT26 tumor-bearing, inflamed and healthy lung ECS. Rates are slopes of linear trace regions (mean ± s.e.m.; healthy and CT26, n = 7; inflamed, n = 3). Two-way ANOVA with Dunnett’s test. P values are provided in Supplementary Table 5. d, DNLGRAF cleavage with protease class inhibitors (150 µM; mean, n = 2). e, Comparison of DNRGRAF versus DNLGRAF (same DNLGRAF data as in c; mean ± s.e.m., n = 7). P values are provided in Supplementary Table 5. Underlined letters highlight the positions at which the two substrates differ (position 3, L versus R). f, Alanine scan of DNLGRAF on-yeast cleavage after 24-hour CT26 ECS treatment (normalized mean ± s.d., n = 3; one-way ANOVA, P < 0.05; Supplementary Table 5). g, Mass spectrometry of CT26 ECS, intact DNLGRAF and DNLGRAF after CT26 ECS treatment showing three cleavage fragments with corresponding mass peaks. For all panels, significance is indicated with asterisk corresponding to ****P < 0.0001, ***P < 0.001, **P < 0.01 and *P < 0.05. Underlined ‘A’ highlight the positions at which the alanine mutations were made. FRET, fluorescence resonance energy transfer; MW, molecular weight; RFU, relative fluorescence units. Graphics in f created in BioRender; Lord, A. https://biorender.com/5uvic7l (2026).
We next sought to identify which tumor-associated protease families contribute to the substrate cleavage. Transcriptomic analysis of patient samples70 revealed upregulation of proteases from multiple families in colorectal adenocarcinoma and skin cutaneous melanoma compared to respective healthy tissues (Supplementary Fig. 5a). Comparison of dysregulation in these two cancer types highlights both shared upregulated proteases, including GZMB, MMP1, MMP9 and unique signatures, including upregulation of MMP3 and MMP7 in colorectal cancer. We assessed the cleavage specificity of the substrates using recombinant catalytic domains of MMP1, MMP2, MMP3, MMP7, MMP8, MMP9, MMP10, MMP11, MMP12, MMP13 and MMP14 (Extended Data Fig. 7b). As expected, PVPLSLVM was cleaved by various MMPs. By contrast, three of the PSurf-identified substrates—DNLGRAF, PFLYLFG and AYGRLYR—were not cleaved by any of the tested MMPs, nor by serine proteases upregulated in colorectal cancer, including FAP14, GZMB69 and PCSK9 (ref. 71), nor by the cysteine protease CTSB—enzymes that have frequently been targeted in recombinant enzyme-based substrate discovery. Among the four new substrates, PARMLHI was the only sequence partially cleaved by certain MMPs (Extended Data Fig. 7b). The discovery of these substrates thus expanded the repertoire of tumor-associated protease sensing beyond the conventional set of enzymes.
To determine which protease families contribute to substrate cleavage, we profiled probe sensitivity using a panel of protease inhibitors, each tested at 150 µM. This analysis did not indicate selective inhibition by a single protease or protease class. Instead, cleavage was most strongly reduced by inhibitors targeting serine and metalloprotease families, particularly when applied in combination (Fig. 4d, Extended Data Fig. 7c and Supplementary Fig. 5b). Specifically, cleavage of DNLGRAF and AYGRLYR was strongly inhibited by the broad-spectrum serine and metalloprotease inhibitor epigallocatechin gallate (EGCG) and by the serine protease inhibitor nafamostat while showing minimal response to cysteine protease inhibitors. Dose–response analysis across multiple inhibitor concentrations yielded half-maximal inhibitory concentration (IC50) values consistent with previously reported ranges (19 µM for nafamostat and 57 µM for EGCG) (Supplementary Fig. 5b)72,73. PARMLHI, consistent with its partial cleavage by MMPs (Extended Data Fig. 7b), was inhibited by MMP inhibitors marimastat and phosphoramidon. By contrast, PFLYLFG was more strongly inhibited by cysteine protease inhibitors, indicating involvement of proprotein convertases and suggesting distinct protease sensitivity compared to the other substrates. These findings indicate that substrates identified by PSurf using tissue-derived samples are responsive to multiple tumor-associated proteases and represent distinct classes of protease-reactive probes compared to conventional MMP substrates.
To investigate how amino acid composition affects substrate sensitivity, we compared substrate DNLGRAF to a substrate derived from the most enriched amino acid at each position: DNRGRAF (Extended Data Fig. 5b). Notably, DNRGRAF differs from the DNLGRAF sequence by only a single residue at the position 3 (R to L) yet showed a substantially reduced ability to differentiate disease states (Fig. 4e). To further investigate the contribution of individual positions, we performed an alanine scan on the DNLGRAF substrate. Each of the six non-alanine residues was individually substituted with alanine, and cleavage activity by CT26 ECS was assessed using yeast display (Fig. 4f). All alanine variants exhibited reduced cleavage compared to the original sequence. Particularly, substitutions at positions 3, 4 and 5 (LGR) resulted in the most pronounced decreases. These findings indicate that efficient substrate activity depends on the precise sequence context rather than on only motif enrichment.
Lastly, we used matrix-assisted laser desorption ionization–time of flight mass spectrometry (MALDI–TOF MS) to identify specific cleavage sites. The untreated probe showed peaks corresponding to the full-length probe and a species lacking its fluorophore (Fig. 4g and Extended Data Fig. 7d). After incubation with CT26 ECS, three additional peaks emerged, corresponding to cleavage among residues L/G, G/R and A/F (Supplementary Table 4). To confirm that these fragments resulted from tissue-derived activity, we monitored the accumulation of higher-mass fragments (>1,000 Da) over time. The progressive increase in fragmented signal supported time-dependent proteolysis (Extended Data Fig. 7d,e). These results indicated that the DNLGRAF substrate undergoes cleavage at multiple specific sites.
These biochemical characterizations demonstrate that PSurf-identified substrates undergo preferential activation in tumor tissue, that protease recognition and cleavage depend on the precise sequence context of the substrate and that multiple protease families contribute to substrate activation and may cleave at multiple sites.
Tumor-activated probe identifies metastases with spatial resolutionTo spatially detect the proteolytic activity in tissues, we designed a probe using the DNLGRAF substrate (MBvivo-DNLGRAF) that enables tissue retention of the probe upon cleavage through electrostatic interactions with a nearby cell membrane only after proteolytic activation (Fig. 5a)74. We first examined the tumor-specific activation of this membrane-binding probe in vivo by comparing retention of the probe in the lungs after intravenous administration. We administered this probe to mice with and without CT26 lung metastases and imaged the lungs ex vivo. The probe demonstrated increased activation in tumor-bearing lungs (Fig. 5b,c), and probe signal correlated with the tumor burden as quantified by tumor autofluorescence (Fig. 5d).
Fig. 5: New substrate-based spatial probe delineates CT26 metastases in whole tissue sections. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, Schematic of MB-DNLGRAF, a membrane-binding fluorogenic probe that emits fluorescence after proteolytic cleavage of the DNLGRAF substrate. b, Schematic of administration of MBvivo-DNLGRAF in mice with and without CT26 lung metastases and fluorescence images of the lungs ex vivo. c, Bar graph showing MFI of MBvivo-DNLGRAF in the lungs with and without CT26 metastases. Data represent the mean ± s.e.m. of n = 8 animals for CT26 and n = 7 for healthy. Unpaired two-tailed t-test was used, **P = 0.0026. d, Correlation analysis of tumor autofluorescence (left) or lung weight (right) with MBvivo-DNLGRAF fluorescence showing a positive correlation in healthy lungs (gray) and CT26 lungs (orange). e, Representative images from two experiments of mouse lung tissue with CT26 metastases stained with hematoxylin and eosin, anti-CA9 or anti-CD31 antibodies or the MB-DNLGRAF probe with or without a protease inhibitor cocktail. Hematoxylin (blue), eosin (red), CA9 (red), CD31 (vasculature, cyan), MB-DNLGRAF (orange). The dashed white lines designate the boundaries of tumor nodules in the lung as identified by H&E in the left-most panel. L, healthy lung; T, tumor nodule.
We further applied this technique to stain CT26 lung metastasis tissue sections using a modified probe format based on the DNLGRAF substrate that enables both fluorescence activation and local membrane insertion upon cleavage (MB-DNLGRAF). DNLGRAF was fused to a membrane-binding peptide75, with a fluorophore at the N terminus and a quencher at the C terminus, enabling cleavage-dependent fluorescence activation. Cleavage by tissue proteases separates the fluorophore from the quencher, leading to dequenching and restoration of fluorescence (Fig. 5e). Hematoxylin and eosin staining identified metastatic lesions, and regions of interest (ROIs) were used to correlate hematoxylin signal with probe fluorescence. MB-DNLGRAF fluorescence showed a strong positive linear correlation with hematoxylin staining (R2 = 0.87) (Extended Data Fig. 8a and Supplementary Fig. 6). MFI was 2.2-fold higher in tumor lesions compared to healthy lung regions (Extended Data Fig. 8b). To confirm protease-dependent probe activation, we applied a cocktail of inhibitors identified from the protease inhibitors screen (Extended Data Fig. 7c) to tissue sections, which reduced tumor-associated fluorescence by 67.3% (Extended Data Fig. 8b). This result demonstrates that probe activation in the tumor is proteolysis dependent.
This probe format was also validated in solution using ECS from healthy lungs and CT26 metastasis-bearing lungs. Consistently, cleavage rates were significantly higher in tumor ECS compared to healthy tissue, and activity was markedly reduced in the presence of inhibitors (Extended Data Fig. 8c,d).
These results demonstrate that the PSurf-identified substrate DNLGRAF undergoes tumor-specific protease cleavage in vivo, in solution and in ex vivo tissue slices, enabling spatial detection of proteolysis activities.
In vivo protease-activated biosensors enable disease classification via urinary readoutWe next leveraged the tumor-activated protease substrates to engineer in vivo biosensors for biofluidic tumor detection. In a modular sensor panel, each of the four PSurf-identified substrates (cleavable linkers) is individually attached to an in vivo trafficking scaffold (for example, nanobody and nanoparticle) and encoded with a chemically stabilized, single-stranded oligonucleotide that enables multiplexing and converts local TME protease activity into a measurable and amplifiable urinary signal (Fig.
Comments (0)