
Remember me
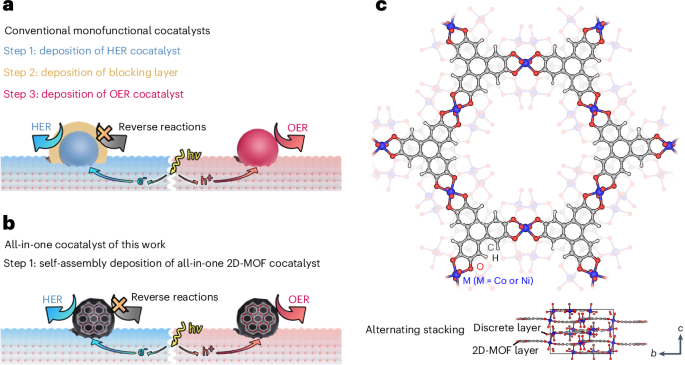
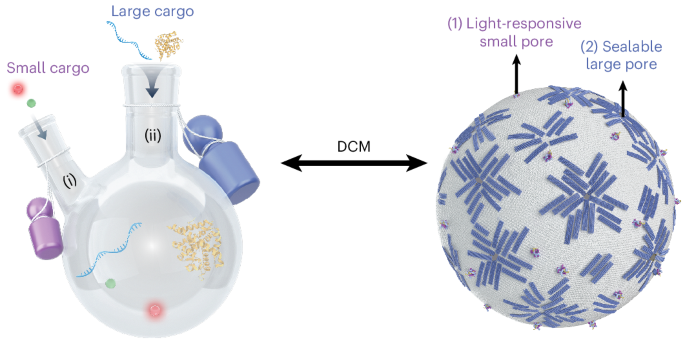
The DCM consists of a GUV (1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC; 0.05 mol% Atto655-DOPE)) as the reactor container, with dynamic SPs and LPs serving as the two necks (Fig. 1a). The light-responsive SPs form the first neck. The SP is a barrel-shaped structure (~12.5 nm in height, ~2 nm inner diameter) with an azobenzene-modified lid strand, allowing reversible light-controlled opening and closing (Fig. 1a(i) and Supplementary Fig. 1). It permits diffusion of small molecules on a minute timescale, as previously characterized34. This lid can open and close the SP in response to light. The as-fabricated SPs are in the closed state. UV illumination (λ = 365 nm) switches the azobenzene from its trans to cis state, opening the pore, while visible light (λ = 450 nm) reverts the azobenzene to its trans state, closing the pore. Upon insertion into the GUV membrane via cholesterol anchors, SPs enable light-gated transport of small molecular cargo across the membrane. The confocal fluorescence images in Fig. 1b demonstrate that UV illumination triggers the opening of SPs, facilitating the influx of sulforhodamine B (SRB) into the GUV. The normalized fluorescence intensity difference (Iout − Iin)/Iout (ref. 44), which quantifies the disparity between the fluorescence intensity inside the GUV (Iin) and in the surrounding bulk solution (Iout), reveals a sharp contrast between the open (SP1) and closed (SP0) states of the light-gated SPs (Fig. 1b). The few outliers observed in SRB influx arise from the intrinsic heterogeneity of GUVs and stochastic SP incorporation. However, the overall trend remains robust across large-population statistical analyses. A control experiment confirms that UV exposure does not affect the permeability of bare GUVs (Supplementary Fig. 2). In addition, statically open (sta-SP1) and closed (sta-SP0) pores, identical in structure but lacking the azobenzene modification, are assembled and examined in the influx experiment (Supplementary Fig. 3). The normalized fluorescence intensity differences of sta-SP1 and sta-SP0 are comparable to those of SP1 and SP0, respectively, confirming the effective gating performance of SPs regulated by light. The light-controlled closure of SPs is further verified by fluorescence recovery after photobleaching (FRAP). Following visible light (λ = 450 nm) illumination, photobleaching of SRB within the GUV leads to an immediate drop in fluorescence intensity. No fluorescence recovery over time is observed (Supplementary Fig. 4), demonstrating the switching reversibility of SPs. In addition to fluorophores, other small molecules essential for cellular activities, such as ions, can also be transported into GUVs via SPs on demand, as demonstrated in the Ca2+ influx experiment (Supplementary Fig. 5).
Fig. 1: DCM. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, A schematic of the DCM, where two dynamic, DNA-based pores synergistically facilitate molecular transport across the GUV membrane with high spatiotemporal control. One neck is represented by light-responsive SPs. The SP featuring an azobenzene-modified lid strand switches between open and closed states via azobenzene photoisomerization under UV and visible light, respectively. The other neck is represented by sealable LPs, formed through the self-arrangement of reconfigurable DRs on the GUV membrane. b, SP transitions between its open (SP1) and closed (SP0) states under UV and visible light illumination, respectively, enabling the transport of small molecules, such as SRB across the GUV membrane. (Iout − Iin)/Iout measurements of the SRB influx are shown for SP0 and SP1. Data represent the mean ± s.d. from three independent experiments. n = 110 in both cases. c, LP formation is induced via a signalling pathway identified using a Cy5-ssDNA (~9 kDa) influx assay. Introducing unlocking strands initiates the transition from s-DRs to e-DRs, leading to GUV deformations. Insertion of SP0 and its subsequent opening (SP1) by UV light triggers GUV shape recovery, leading to LP formation (LP1).
Recently, we have demonstrated that DNA origami rafts (DRs) can undergo reversible conformational changes on GUV membranes, switching from a square shape (s-DR, 70.8 nm × 55 nm, aspect ratio ~1.3) to an elongated rectangle shape (e-DR, 190 nm × 20 nm, aspect ratio ~9.5) upon the addition of DNA fuels45 (Fig. 1a(ii)). This self-arrangement of DRs from an isotropic to a short-range local order (Supplementary Fig. 6) generates steric pressure, inducing membrane bending. The localized deformation collectively translates into stress, driving large-scale morphological changes of the GUV45. Assisted by the statically open protein pore OmpF, the locally ordered DRs perforate the membrane during GUV shape recovery, forming large membrane pores. In this work, we extend the generality of this working mechanism by replacing protein pores with light-responsive SPs to induce DNA-based, sealable LPs mediated by GUV membrane dynamics. This enables the creation of the DCM entirely from synthetic building blocks, which can be independently controlled to facilitate programmable and dynamic coordination, thereby driving emergent complexity in synthetic reaction systems.
As shown in Fig. 1c(i) and Supplementary Fig. 7a, when only s-DRs (Atto488 labelled) are bound to the membrane, the GUV maintains a spherical shape. The addition of unlocking strands triggers the transition from s-DRs to e-DRs, resulting in prominent membrane deformations (Fig. 1c(ii) and Supplementary Fig. 8). When SPs in their closed state (SP0) are integrated into the GUV membrane, the deformations persist and no apparent influx of ssDNA (~9 kDa, Cy5 labelled) is observed (Fig. 1c(iii) and Supplementary Fig. 7b). Upon UV illumination, the opening of SPs (SP1) allows the exchange of small solutes (for example, sucrose, Na+, Mg2+ and Cl−) across the membrane, rapidly restoring osmotic balance. This process is further supported by the observed Ca2+ influx detected using the Ca2+-responsive fluorophore Fluo-8 (Supplementary Fig. 9). During this process, the GUV gradually returns to its spherical shape, yet no influx of ssDNA is detected (Fig. 1c(iv)). Only after the GUV fully recovers its spherical shape does the transport of ssDNA start, indicating the formation of LPs (LP1; Fig. 1c(v) and Supplementary Figs. 7c and 8) through interactions between the self-arranged e-DRs and the GUV membrane. The influx reaches equilibrium after ~30 min (Supplementary Fig. 10). These LPs continue to facilitate transmembrane transport of ssDNA even after 2 days (Supplementary Fig. 11), demonstrating their non-transient nature and long-term stability. Also, s-DR-bound GUVs integrated solely with light-responsive SPs in the open state (SP1) show no influx of ssDNA due to size exclusion (Supplementary Fig. 12). e-DR-bound GUVs with light-responsive SPs in the closed state (SP0) exhibit neither shape recovery nor ssDNA influx (Supplementary Fig. 13). Together with a theoretical framework that qualitatively elucidates the underlying mechanism (Supplementary Fig. 14 and the discussion immediately following), these experiments demonstrate that the LP formation is not random but is tightly regulated by a defined signalling pathway. This pathway during DCM formation involves the input of unlocking strands to trigger the transition from s-DRs to e-DRs, GUV deformations, the opening of SPs (SP1) by UV light, recovery of the GUV’s shape and eventually, large pore formation (LP1)45. Thus, the indirect interaction between SPs and LPs (Supplementary Fig. 15), mediated by the membrane dynamics of the GUV, leads to the creation of the DCM equipped with two fully synthetic, dynamic necks for orchestrated molecular delivery.
Dynamic control over the DCM permeabilityThe dynamic interplay between SPs and LPs enables the DCM to adopt four distinct states based on their on/off configurations (Fig. 2a). Visible light illumination closes SPs, transitioning the DCM from SP1-LP1 to SP0-LP1. Subsequent addition of locking strands reconfigures e-DRs back to square DNA rafts (R-s-DRs), sealing LPs and resulting in SP0-LP0. Alternatively, the DCM transitions from SP1-LP1 to SP0-LP0 by first adding locking strands to reach SP1-LP0, followed by visible light illumination. To evaluate the sealing efficiency of LPs, FRAP experiments are conducted at SP0-LP0 and SP0-LP1, respectively, for direct comparison. At SP0-LP0, photobleaching of Cy5-ssDNA within the GUV results in an immediate decrease in fluorescence intensity without recovery over time (Fig. 2b, grey curve, and Supplementary Fig. 16a), confirming the successful closure of LPs. By contrast, at SP0-LP1, fluorescence recovers rapidly after photobleaching (Fig. 2b, blue curve, and Supplementary Fig. 16b). Beyond ssDNA, other large molecular cargo, such as fluorescein isothiocyanate (FITC)–dextran molecules (∼20 kDa) and green fluorescent protein (GFP, ∼27 kDa) can also be efficiently transported through LPs (Fig. 2c and Supplementary Fig. 17). In addition, the dimension of LPs can be determined through influx assays using molecules of varying molecular weights44 (Fig. 2d and Supplementary Figs. 18 and 19). The population distribution centres at 0.56 for 70 kDa FITC–dextran, suggesting that the dimension of LPs is ~15 nm based on the hydrodynamic diameter of the dextran molecule46. Consistently, the DCMs also enable size-selective efflux of encapsulated molecules, demonstrating bidirectional and regulated transport (Supplementary Fig. 20).
Fig. 2: Dynamic control over the DCM permeability. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, The on/off configurations of SPs and LPs establish four distinct states of the DCM: SP1-LP1, SP0-LP1, SP1-LP0 and SP0-LP0, where ‘1’ and ‘0’ denote on and off states, respectively. b, The sealing efficiency of LPs. At SP0-LP0, photobleaching of ssDNA-Cy5 within the GUV causes a sharp fluorescence intensity decrease without recovery (grey), confirming successful LP closure. Conversely, at SP0-LP1, fluorescence recovers rapidly after photobleaching (blue). Data are presented as mean ± s.d. (n = 3 for both cases). c, The efficient transport of large cargo, including ssDNA (9 kDa), dextran (20 kDa) and GFP (27 kDa), is demonstrated via influx assays, measured as (Iout − Iin)/Iout. Inset: scale bars, 5 μm. Data represent mean ± s.d. from three independent experiments. n = 97, 88 and 96 for ssDNA, dextran and GFP, respectively. d, The dimension estimation of LPs based on influx assays using molecules of varying molecular weights. Data represent mean ± s.d. from three independent experiments. n = 114, 114, 77, 112, 110, 114, 114, 117, 120 and 114, for Cy3, 5 kDa ssDNA, 9 kDa ssDNA, 20 kDa dextran, GFP, 40 kDa dextran, 70 kDa dextran, 150 kDa dextran, 500 kDa dextran and 2,000 kDa dextran, respectively. e, The dynamic interplay between SPs and LPs allows precise temporal control over DCM permeability, facilitating orchestrated cargo transport.
A key feature of the DCM is its dynamically controllable permeability through the coordination of SPs and LPs. To demonstrate this functionality, influx assays are performed with Cy5-ssDNA and SRB coexisting in the exterior of GUVs (Fig. 2e and Supplementary Fig. 21), while e-DRs are labelled with Atto488 to define membrane boundaries. When the DCM is at SP1-LP1, Cy5-ssDNA is transported via LPs, while SRB diffuses through both LPs and SPs. At SP0-LP1, where SPs are closed upon visible light exposure and LPs remain open, both Cy5-ssDNA and SRB are transported into the GUV lumen through LPs. At SP1-LP0, where SPs are opened by UV illumination and LPs are sealed by reconfiguration from e-DRs to R-s-DRs, only SRB influx occurs because size exclusion prevents Cy5-ssDNA entry. Finally, at SP0-LP0, no influx is observed. In addition, a detachable raft strategy is developed to enable reliable system resetting and LP re-opening through toehold-mediated raft detachment, membrane resealing and reconfiguration upon resupply of s-DRs (Supplementary Figs. 22 and 23). Furthermore, the DCM platform exhibits stability and functional compatibility under physiologically relevant conditions47,48 (Supplementary Fig. 24).
In typical GUV-confined reactions, large reactants are often pre-encapsulated during GUV formation or mechanically injected. Here, the coordination between SPs and LPs enables sequential, on-demand transport of different reactants into DCMs, as exemplified by the implementation of an enzyme cascade. Enzyme cascades are a hallmark of biochemical organization in living systems, where sequential reactions are orchestrated with spatial and temporal precision. A GOx–myoglobin cascade is established within the DCM, regulated through the stepwise transport of distinct reactants (Supplementary Fig. 25). The realization of this cascade within a membrane-confined environment highlights that the DCM can go beyond single-step conversions to achieve programmable, pathway-like biochemical processing. In the following sections, we further showcase how these unique features facilitate biochemical reactions inside DCMs with programmable transport and precise spatiotemporal control.
Actin polymerization and bundling within the DCMActin polymerization and bundling are vital biological processes that contribute to cell shape, motility and intracellular transport49,50. G-actin, a globular protein, polymerizes into long, filamentous structures (F-actin) within cells51,52. These filaments are then organized into bundles by crosslinking proteins such as fascin53,54,55, which enhances their structural stability and mechanical strength (Supplementary Fig. 26).
The DCM provides a controlled, cell-sized environment to reconstruct actin networks through on-demand delivery of reactants (Fig. 3a). DRs and G-actin are labelled with Atto488 and rhodamine, respectively. During the transition from s-DRs to e-DRs, GUV deformation occurs, followed by SP activation with UV light and subsequent GUV shape recovery. This signalling pathway enables the formation of LPs, facilitating the influx of G-actin (~42 kDa) and its uniform distribution within the lumen (Supplementary Fig. 27). After verifying G-actin transport, unlabelled G-actin is used in actin polymerization and bundling experiments with Acti-stain 488 phalloidin employed for filament staining and visualization (Fig. 3b). Initially, the DCM lumen appears dark at SP1-LP1 (Fig. 3b(i)). G-actin is then transported through LPs, while Acti-stain 488 phalloidin is added. The lumen exhibits homogeneous fluorescence (Fig. 3b(ii)). ATP is subsequently delivered via both SPs and LPs, binding to G-actin to initiate polymerization. The formation of elongated, filamentous structures confirms F-actin assembly (Fig. 3b(iii) and Supplementary Fig. 28). Finally, fascin (~55 kDa) is introduced into the DCM through LPs, crosslinking the polymerized actin filaments. This crosslinking creates densely packed actin bundles and interconnected networks, emulating the organizational complexity of cellular actin cytoskeleton (Fig. 3b(iv) and Supplementary Fig. 29).
Fig. 3: Actin polymerization and bundling within the DCM. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, A schematic of sequential transport of reactants, enabling actin polymerization and bundling within the DCM. b, Intermediate steps and corresponding confocal fluorescence images of the DCM: (i) DCM is initially at SP1-LP1; (ii) after G-actin (~42 kDa) transport through LPs, the lumen exhibits homogeneous fluorescence; (iii) ATP is delivered via both SPs and LPs, binding to G-actin to initiate polymerization; and (iv) fascin (~55 kDa) is introduced into the DCM through LPs, crosslinking the polymerized actin filaments. Lipids (Atto655), filament staining and visualization (Atto488-phalloidin).
Cell-free RNA transcription within the DCMRNA transcription is a fundamental process of cellular gene expression and regulation56,57,58. To demonstrate the versatility of our platform, we next implement cell-free RNA transcription within the DCM (Fig. 4a). This involves the sequential delivery of a transcription mixture (TX mix) and a fluorescence indicator, recapitulating key aspects of transcriptional regulation.
Fig. 4: Spatiotemporally controlled cell-free RNA transcription within the DCM. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, A schematic of Spinach RNA aptamer synthesis and fluorescence visualization in a cell-free transcription (TX) system regulated by spatiotemporally regulated transport of reactants. b, The stepwise workflow and corresponding confocal fluorescence images: (i and ii) DNA templates and TX mix are introduced into DCMs pre-encapsulating T7 RNA polymerase; (iii) both LPs and SPs are closed to create a confined environment and prevent leakage after removal of exterior templates; and (iv) SPs are subsequently re-opened to allow DFHBI influx, enabling fluorescence visualization of RNA production. Green, DRs and Spinach–DFHBI complex; red, lipids and Cy5-labelled DNA template.
As illustrated in Fig. 4b, the Spinach RNA aptamer is employed as a model reporter, emitting fluorescence upon binding to the fluorogen 3,5-difluoro-4-hydroxybenzylidene imidazolinone (DFHBI; ~252 Da), thus providing a convenient optical readout58,59. T7 RNA polymerase (~99 kDa) is pre-encapsulated in GUVs during preparation and remain confined after DCM formation in the SP1-LP1 state. The TX mix, containing Cy5-labelled DNA templates (~35 kDa), NTPs and transcription buffer, is introduced externally and transported into the DCM. Both LPs and SPs are subsequently closed to create a sealed, cell-like compartment and prevent leakage after removal of residual exterior reagents. Following incubation at 37 °C for 2 h, SPs are re-opened by UV illumination to allow DFHBI influx. Clear green fluorescence from the Spinach–DFHBI complex is observed within the DCM lumen, confirming RNA production. These results demonstrate that the DCM can achieve controlled, cell-free gene expression within a confined, programmable synthetic compartment. Furthermore, large field-of-view confocal fluorescence images (Supplementary Fig. 30) illustrate the high functional fidelity and reliability of our system.
Confined synthesis of 3D DNA crystals within the DCMHierarchical DNA self-assembly, where small DNA motifs form larger structures through multistep processes, substantially expands the complexity and diversity of DNA-based materials17. In 2009, Seeman and Mao introduced the rational design of a 3D DNA crystal based on the tensegrity triangle, creating a well-ordered macromolecular 3D crystalline lattice60. Later, they improved sticky-end cohesion via 5′-phosphorylation, effectively promoting DNA crystal nucleation and growth61,62,63.
Traditionally, DNA crystals are synthesized in bulk solution drops, requiring prolonged evaporation or slow annealing60,61,62,63,64,65,66. However, the confined growth of DNA crystals remains largely unexplored, despite its promising scientific and technical potential. For instance, integrating DNA crystals into synthetic cells could advance their functional capabilities by providing scaffolds to spatially organize biomolecules and enzymes with high precision. We demonstrate the synthesis of 3D DNA crystals within cell-sized confinements. As a model system, we use Seeman and Mao’s 3D rhombohedral DNA crystals (Fig. 5a), where DNA triangle motifs connect via sticky-end cohesion modified by 5′-phosphorylation63. This crystallization process is highly sensitive to assembly conditions, requiring precise control over DNA triangle motif concentration and buffer parameters, such as ionic strength and sucrose concentration. Optimized buffer conditions of 50 mM sucrose (Supplementary Fig. 31) and 10 mM Mg2+ (Supplementary Fig. 32) are first identified for crystal growth at DNA triangle motif concentrations exceeding 10 µM (Supplementary Fig. 33). A lower Mg2+ concentration of 5 mM proves insufficient to induce crystallization.
Fig. 5: Confined synthesis of 3D DNA crystals within the DCM. The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.a, A schematic of 3D DNA crystallization within cell-sized confinements. b, Intermediate steps and corresponding confocal fluorescence images of the DCM: (i) DCM is initially at SP1-LP1; (ii) DNA triangle motifs are introduced through LPs into the DCM; (iii) LPs are sealed by adding locking strands; and (iv) Mg2+ is incrementally supplemented into the DCM through SPs to induce DNA crystallization. Green, DRs; red, Cy5-labelled DNA motifs. c,d, Bright-field image (c) and z-stack 3D visualization (d) of a DCM containing a single DNA crystal. e, Statistical analysis of the number of DNA crystals formed in individual DCMs. n = 44. f, Birefringent properties of the DNA crystals under crossed polarized light. The alternating bright and dark appearances of the two DNA crystals under rotating polarizers confirm the presence of distinct crystalline axes. The orientations of the polarizer (P, dashed arrow) and analyser (A, solid arrow) are indicated by arrows.
To synthesize DNA crystals, DCMs are prepared with both SP and LPs open (SP1-LP1; Fig. 5b(i)). DNA triangle motifs, pre-assembled by annealing three composing DNA strands, are introduced into the DCMs through LPs in the presence of 5 mM Mg2+. Cy5-labelled DNA triangle motifs distribute homogeneously within the DCM lumen (Fig. 5b(ii)). LPs are then sealed by adding locking strands, converting e-DRs to R-s-DRs (Fig. 5b(iii)). This sealing step confines the DNA motifs within the DCM, preventing efflux during subsequent steps. Mg2+ is incrementally supplemented into the DCM through SPs to achieve a final concentration of 10 mM. Stepwise Mg2+ supplementation is crucial because directly introducing 10 mM Mg2+ at the start would cause crystallization primarily outside the DCM, where reactants are abundant. Controlled Mg2+ delivery through SPs, combined with sealed LPs, ensures effective nucleation and crystal growth, as evidenced by the single-crystal formation within the DCM (Fig. 5b(iv)). The DNA crystals are formed inside the DCMs following a 2-day incubation at 22 °C. Figure 5c,d shows the bright-field and 3D reconstruction (Supplementary Fig. 34) images of a DCM containing a single DNA crystal with defined facets, characteristic of its crystalline nature.
DNA crystallization within DCMs demonstrates notable confinement effects. Confinement predominantly promotes nucleation at a single site (72.7%, n = 44; Fig. 5e), suppressing multinucleation events. Once nucleation initiates, it tends to dominate the growth process, outcompeting other potential sites owing to the depletion of DNA motifs in the confined space. As reactants are sequentially delivered into the DCM, the first site where Mg2+ reaches a critical nucleation concentration starts crystal growth, reducing supersaturation to levels insufficient for additional nucleation events. Furthermore, crystallization preferentially occurs near the DCM’s inner surfaces (Supplementary Fig. 35), probably because these surfaces provide favourable nucleation sites. As Mg2+ is introduced through SPs in the membrane, its concentration is probably higher near the inner surface, driving crystallization in that region. In addition, the confined volume of the DCM limits crystal growth, favouring boundary-aligned development. The crystalline nature of the 3D DNA crystals is validated through birefringence analysis using crossed polarizers. In a DCM containing two DNA crystals (Fig. 5f), the alternating bright and dark appearances under rotating polarizers confirm the presence of distinct crystalline axes (also see Supplementary Fig. 36).
Comments (0)