
Remember me
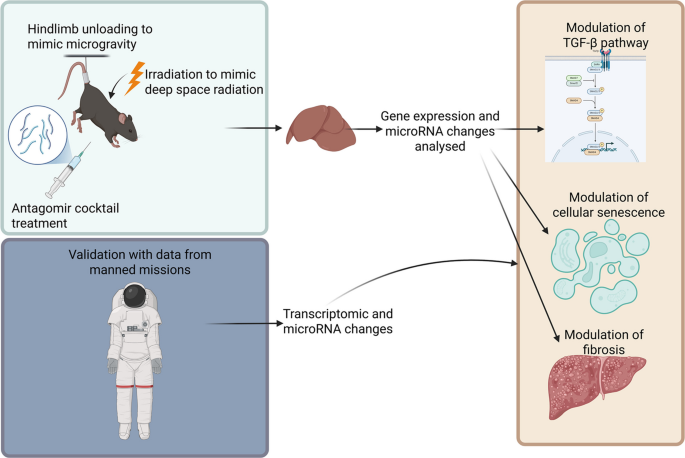
To evaluate the potential of the space environment to alter miRNA and mRNA expression patterns in the liver, RT-PCR was conducted on tissue extracted from the liver of mice exposed to 0.5 Gy galactic cosmic radiation (GCR), 1.0 Gy solar particle event (SPE) irradiation, and hindlimb unloading (HU) to simulate microgravity (Fig. 1). We found that hindlimb unloading alone significantly altered the expression of many miRNAs involved in the TGF-β pathway, with increased expression of miR-16-5p, miR-125b-5p, miR-21-5p, miR-27a-3p, and miR-148a-3p (Fig. 2A). However, these changes in miRNA expression caused by simulated microgravity were not correlated with any significant changes in mRNA expression.
Fig. 1
Summary timeline of animal experiments. Female 15-week-old C57BI/6J wild-type mice experienced 14 days of normal loading (NL) or hindlimb unloading (HU), using the Morey-Holton method (30° head-down) to mimic microgravity conditions. Antagonists to the miRNAs miR-16-5p, miR-125b-5p, and let-7a-5p were administered by intraperitoneal injection at a dose of 10mg/kg each (total dose 30 mg/kg) every 3 days, starting 1 day before HU setup and ending 1 day before irradiation. 0.5 Gy GCR and 1.0 Gy SPE irradiation were delivered on the 13th day. Twenty-four hours after irradiation, mice were euthanized, and livers were dissected and snap frozen
Fig. 2
Heatmap illustrating the overall expression profile of the 8 miRNAs and 15 mRNAs studied in response to conditions including: A microgravity/hindlimb unloading (HU), B 0.5 Gy galactic cosmic radiation (GCR), and C 1.0 Gy solar particle event (SPE) irradiation with and without antagomir treatment. The studied mRNAs are grouped by pathway: TGF-β pathway, fibrosis, senescence/cell cycle, and inflammation. Colors represent the −ΔΔCt value (log2(FC)) from downregulated in blue to upregulated in red. p-values less than 0.05 of the treatment group compared to the control Sham Ag- NL group using both paired t-test (#) and a more stringent multi-factor ANOVA (*) were shown
Furthermore, we evaluated the effects of GCR and SPE irradiation on expression and found that changes to both miRNA and mRNA expression profiles largely differed depending on the type of radiation given (Fig. 2B and C). Notably, SPE irradiation significantly altered the expression of several miRNAs involved in the TGF-β pathway, with downregulation of let-7a-5p, let-7b-5p, miR-21-5p, and upregulation of miR-27a-3p (Fig. 2C). In combination with microgravity, SPE irradiation also decreased the expression of miR-125b-5p. Interestingly, GCR irradiation alone caused no significant shifts in miRNA expression, but when combined with microgravity increased the expression of miR-125b-5p and miR-181b-5p and decreased the expression of miR-27a-3p.
Moreover, the type of radiation given also influenced whether shifts in miRNA expression correlated to observed changes in mRNA expression. Notably, in both the GCR and SPE groups, there was an increased mRNA expression of p21, a gene associated with senescence (Fig. 2B, C). Additionally, SPE irradiation increased expression of Col1a1 and Col4a1, two fibrosis-associated genes. While none of the downregulated miRNAs observed have been confirmed as direct targets for p21, downregulation of miR-125b, miR-16, let-7a, and miR-148a has all been associated with senescent cells [33]. Furthermore, let-7a, miR-148a, and miR-125b have also been shown to influence liver fibrosis via the TGF-β pathway; however, none have been confirmed to be direct targets of Col1a1 or Col4a1 expression [34]. These findings suggest that the radiation encountered in space, especially SPE irradiation, represents high risks for hepatic cellular senescence and fibrotic conditions.
Of note, female C57BL/6 J mice were used in this study to maintain consistency with the majority of previous spaceflight and spaceflight-analog studies. This alignment enables direct comparison with existing datasets from NASA’s GeneLab and other spaceflight missions, thereby enhancing translational relevance while minimizing experimental variability under resource constraints.
Antagomirs successfully decrease target miRNA expression modulating the expression profileBecause of their role in TGF-β1 pathway regulation, which has previously been shown to decrease with increasing radiation dose, antagomirs to the miRNAs miR-16-5p, miR-125b-5p, and let-7a were administered prior to radiation exposure to evaluate their ability to modulate transcriptional responses within the TGF-β pathway [29]. Within the non-irradiated group, antagomir (Ag) treatment successfully decreased expression of miR-125b-5p and let-7a-5p in both the NL and HU groups compared to their untreated counterparts (Fig. 2A). Additionally, miR-16-5p expression is significantly decreased in these groups when compared to the Sham HU group (Figs. 2A and 3A). Importantly, antagomir treatment remained effective when exposed to radiation. Compared to the untreated Sham group, Ag treatment decreased the expression of all 3 of its targets in both the GCR and SPE groups. More specifically, within the GCR group, Ag treatment decreased expression of miR-125b-5p and let-7a-5p regardless of the presence of microgravity (Fig. 3B, C). Ag treatment seemed to be least effective in the SPE groups, only decreasing expression of miR-16-5p in combination with hindlimb unloading compared to its untreated SPE NL and HU counterparts (Fig. 3A).
Fig. 3
RT-PCR analysis plots of miRNAs involved in the TGF-β pathway showing miRNA expression fold change by radiation, antagomir treatment, and microgravity. The −∆∆Ct method was used to calculate and normalize the fold change, in which −∆∆Ct was calculated as −∆∆Ct = average (∆Ctcontrol) − ∆Ctsample, and ∆Ct = Cttarget − Cthousekeeping. The housekeeping miRNA was U6, and Sham NL Ag(-) samples were set as control samples. Multiple factor ANOVA was performed to evaluate the partial effect of radiation, antagomir treatment, and microgravity. (A) miR-16-5p, (B) miR-125-5b, (C) let-7a-5p, (D) let-7b-5p, (E) miR-21-5p, (F) miR-27a-3p, (G) miR-148a-3p and (H) miR-181b-5p
By downregulating the expression of its 3 targets, antagomir treatment also influenced the expression of the other TGF-β pathway miRNAs studied. In all three radiation groups, Ag treatment was associated with decreased expression of let-7b-5p and miR-21-5p when compared to the untreated Sham group (Fig. 2). Within the Sham group, Ag treatment seemed to prevent the effects of microgravity on miR-181b-5p, significantly decreasing its expression from slightly upregulated in the untreated HU group to significantly downregulated in the treated HU group (Fig. 3H). Within the hindlimb unloaded GCR group, Ag treatment had a similar effect on miR-181b-5p, decreasing its expression from significantly upregulated to significantly downregulated (Figs. 2 and 3H). Remarkably, Ag treatment had opposite effects on miR-27a-3p expression depending on the type of radiation given. In the GCR group, Ag treatment prevented the combined effects of GCR and microgravity, increasing its expression from significantly downregulated back to baseline (Fig. 2A, B). We have shown that regulation of this miRNA signature is most likely specific for GCR exposure, and it is probable that there will be other key miRNAs to inhibit for SPE exposure [28]. But despite this fact, we still do see changes occurring as expected when inhibiting these miRNAs under SPE irradiation. Interestingly, Ag treatment decreased expression of miR-27a-3p in the SPE groups, from significantly upregulated back to baseline under normal loading and to significantly downregulated under simulated microgravity (Fig. 2A). Overall, antagomir treatment successfully decreased expression of its targets, offering a varied yet potentially protective effect against the impacts of radiation and microgravity on other miRNAs involved in the TGF-β pathway.
Antagomir treatment modulates TGF-β-associated transcription, senescence, and inflammation-related gene expression, while further increasing fibrosis-associated markers. Based on our compelling results showing the modulation of the expression of genes in response to antagomir treatment—which inhibits 3 key miRNAs involved in the TGF-β pathway—on miRNA expression, we further investigated its influence on the expression of mRNAs involved in the TGF-β pathway, along with the downstream processes of fibrosis, senescence, and inflammation. We found that Ag treatment preserved Tgfb1 expression against the combined influence of SPE and microgravity, increasing its expression from significantly downregulated back to baseline (Fig. 2A, C). Under GCR irradiation, Ag treatment had a wider effect on the TGF-β pathway genes studied, significantly increasing most of their expression from slightly downregulated to significantly upregulated. Decreased let-7b expression, which was observed in the Ag-treated GCR group but not the treated SPE group, has been linked to increased expression of Tgfb1 and Tgfbr1 expression [35]. Therefore, its altered expression might be partially explanatory for the increased expression of genes involved in the TGF-β pathway changes in this group.
Antagomir treatment also seems to shift the transcriptomic profile for genes involved in senescence, the cell cycle, and inflammation. Expression of mtor, a marker associated with aging, was restored from significantly downregulated back to baseline under microgravity in both the GCR and SPE groups (Fig. 2B, C). Previous work shows transfection of a miRNA cocktail containing miR-16-5p significantly decreased MTOR expression in the liver, so perhaps its inhibition by the current antagomir treatment is partially responsible for the observed increase [36]. Furthermore, Ag treatment maintained expression of Ccnd1, Il6, and Cxcl1—increasing their expression from significantly downregulated under GCR back to baseline (Fig. 2B). Previous findings indicate that CCND1 protein levels decrease with aging in the livers of mice, so by restoring Ccnd1 expression, the current treatment may shift gene expression patterns associated with cellular aging under GCR exposure [36]. Additionally, downregulation of CXCl1 was observed in patients with alcoholic hepatitis; therefore, its restoration may reflect transcriptional changes associated with inflammatory regulation under GCR exposure [37]. Furthermore, Il6 is a direct target of miR-125b; thereby, its inhibition could play a role in the observed expression changes [38]. Interestingly, Ag treatment also significantly upregulated expression of anti-inflammatory Il10 in all radiation groups.
Unfortunately, antagomir treatment also increased expression of both studied fibrotic markers. Within the GCR group, Ag treatment significantly increased expression of Col1a1 and Col4a1 from slightly upregulated to significantly upregulated (Fig. 2B and Supplemental Fig. S2). Moreover, Col1a1 and Col4a1 expression was significantly upregulated in the SPE group both with and without Ag treatment (Fig. 2C). Previous studies found that a miR-125b mimic restored fibrotic liver damage in CCl4-treated rats, so perhaps its inhibition by antagomir treatment is responsible for the increased fibrotic markers observed in the current study [39]. Overall, antagomir treatment was effective at changing the levels of associated mRNAs, offering transcriptional modulation for genes involved in the TGF-β pathway, senescence, and inflammation, but also exacerbating the increased expression of fibrotic genes.
Correlation analysis reveals miRNA and mRNA profiles function synergistically yet inverselyAs discussed above, inhibition of 3 key miRNAs involved in the TGF-β pathway by antagomir treatment induced widespread changes to both the miRNA and mRNA expression profiles. Therefore, we further explored these relationships by performing pathway analysis using DIANA-TarBase, which reveals that many of the miRNAs whose expression was upregulated by the given antagomir treatment work to inhibit expression of genes involved in the TGF-β pathway (Fig. 4). Spearman correlation analysis was also conducted to further evaluate the relationship between the studied miRNA and mRNA expression profile (Fig. 5). General analysis using data from all samples illustrates an overall trend of positive correlation among the 8 studied miRNAs with 15 significant correlations (p < 0.05), 7 of which are highly significant (p < 0.001) (Fig. 5A) (Table S1). Likewise, the mRNA expression profile appears to function synergistically with 42 significant positive correlations (p < 0.05) among 15 studied mRNAs, 10 of which are highly significant (p < 0.001). Notably, there is a pronounced negative correlation between expression of the studied miRNAs and mRNAs, in which miRNA upregulation was associated with mRNA downregulation, with 31 significant inverse correlations (p < 0.05). Interestingly, the observed negative correlation between miRNA and mRNA expression appears largely specific to GCR and microgravity conditions, with 25 significant inverse correlations (p < 0.05), when compared to only 4 significant inverse correlations (p < 0.05) under SPE and microgravity conditions (Fig. 5B, C) (Table S2). When specifically analyzing the impact of the three miRNAs targeted by the given antagomir treatment, miR-16-5p, miR-21-5p, and let-7a-5p, on the expression of target and downstream mRNAs, the same trend is observed with 8 significant inverse correlations (p < 0.05) under GCR and microgravity conditions, compared to 2 significant inverse correlations under SPE and microgravity conditions. Likewise, their expression is positively correlated with the expression of the other studied miRNAs, with let-7a-5p and miR-16-5p having 5 significant positive correlations and miR-21-5p having 4 across all samples (Fig. 5A).
Fig. 4
miRNAs are involved in the regulation of genes associated with the TGF-β pathway. A Illustration of the TGF-β pathway showing proteins of interest and miRNA targets as predicted using the DIANA-TarBase v8 tool and previous literature findings. Pathway map created with BioRender.com. B Network plot showing connections between all studied miRNAs and mRNAs. Network plot was made using Cytoscape
Fig. 5
Spearman correlation plots showing correlations between all studied miRNAs and mRNAs with a single asterisk representing p = 0.05, two asterisks representing p = 0.01, and three representing p = 0.001. A Correlation plot based on averages of all samples. B Correlation plot based on averages of samples treated with 0.5 Gy GCR irritation and simulated microgravity. C Correlation plot based on averages of samples treated with 1.0 Gy SPE irradiation and simulated microgravity
From mouse tissue to human: translational potential of on-earth mice studies to human astronautsAfter detecting altered expression of markers associated with fibrosis, senescence, and inflammation in mice livers exposed to the simulated space environment which were modulated at the transcriptional level by antagomir countermeasures, we analyzed astronaut data from the National Aeronautics and Space Administration (NASA) Twin Study mission, in which an astronaut was studied during and after 340days spent aboard the International Space Station (ISS), and the Inspiration4 (I4) mission, a 3-day private civilian spaceflight in low Earth orbit (LEO) [40,41,42]. It must be noted that both missions remained in LEO and thus experienced significantly lower doses of radiation than the 0.5 Gy GCR estimated for a flight to Mars and back or the 1.0 Gy SPE irradiation estimated for unshielded exposure to a solar flare that this study’s mice were exposed to. Indeed, the NASA Twin Study astronaut was exposed to 14.634 cGy of irradiation, and the I4 astronauts 0.472 cGy [40, 41]. To further investigate the mRNA expression profile of several genes whose expression was restored with antagomir treatment, RNA-sequencing was performed on blood cell fractions. More specifically, for the Inspiration4 mission, blood samples were collected from the 4 astronauts—2 male and 2 female—preflight, 1-day postflight, 45 days postflight, and 82 days postflight. For the NASA Twin Study, both mRNA and miRNA-sequencing were performed on blood samples collected from a singular 50-year-old male astronaut before, during, and up to 200 days after a 340-day mission on the ISS.
Even with the relative protection of Earth’s magnetosphere in LEO, astronauts experienced expression changes of many of the miRNAs which were targeted by antagomir treatment in mice that are exposed to Mars-mission-equivalent acute high-dose radiation simulations. In the NASA Twin study, 4 blood cell populations were evaluated—CD4 T-cells, CD8 T-cells, CD19 B-cells, and lymphocyte-depleted (LD) cells (Fig. 6). However, overall trends are best represented by the aggregate of all peripheral blood mononuclear cells (PBMC), in which expression of let-7a, miR-125b, miR-21, and miR-148a was upregulated long after return to Earth (R + 103–200). When analyzing the mRNA expression of the same PBMC blood cell fraction, expression of 5 of the 6 studied genes directly involved in the TGF-β pathway (SMAD4, TGFBR2, TGFB1, SMAD2, TGFBR1) was downregulated after lengthy time spent in space, flight day (FD) 181–334, but expression of these genes was largely restored by the first collection taken after return (R + 103) (Fig. 7). Interestingly, mRNA expression varied by cell type with inverse expression patterns observed between the CD19 and LD cell fractions. Data from the I4 mission samples a wider variety of circulating cell types, including PBMCs, B-cells, CD4, CD8, CD14, CD16, dendritic cells, natural killers, and all other T cells (Fig. 8). While expression of many of the studied genes varies between cell type across the 3 post-flight timepoints, slight downregulation of TGFB1 on the first day after return (R + 1) is conserved across every cell type. However, its expression is largely restored by the last sample collection (R + 82). Notably, upregulation of SMAD2 and SMAD4 was recorded 1 day after return in 9 of the 10 cell types. Overall, RNA-seq data from the NASA Twin and I4 studies highlight a pattern of downregulation for several genes in the TGF-β pathway during or shortly after return to Earth, but they are restored upon further time spent on Earth.
Fig. 6
Heatmap showing changes in expression of miRNAs involved in regulation of the TGF-β pathway at various timepoints preflight, during flight, and postflight using data from the NASA Twin Study. Plots generated for various white blood cell types, including CD4, CD8, CD19, CPT (PBMCs), and lymphocyte-depleted (LD). FD indicates flight day number, and R + represents days after return to Earth
Fig. 7
Heatmaps showing changes in expression of mRNAs involved in fibrosis, senescence, and inflammation at various timepoints preflight, during flight, and postflight using data from the NASA Twin Study. Plots generated for various white blood cell types, including CD4, CD8, CD19, CPT (PBMCs), and lymphocyte-depleted (LD). FD indicates flight day number, and R + represents days after return to Earth
Fig. 8
Heatmaps showing changes in expression of mRNAs involved in fibrosis, senescence, and inflammation at various timepoints preflight and postflight using data from the Inspiration4 mission. Plots generated for various white blood cell types, including PBMCs, B, CD4, CD8, CD14, CD16, DC, NK, and all other T cells
To support an association between changing miRNA expression and the expression of mRNAs involved in fibrosis, senescence, and inflammation in astronauts, RNA-seq data from the NASA Twin study for 14 selected miRNAs and 11 mRNAs were analyzed. Among the four studied cell types, significant inverse correlations between 2 of the 3 miRNAs inhibited by antagomir, miR-16 and let-7a, and expression of CXCL1, CCND1, and TGFB1 were noted (Fig. 9). Furthermore, significant inverse relationships between several other miRNAs—whose expression was downregulated synergistically with the 3 direct targets—and other genes involved in the TGF-β pathway, including TGFB1, TGFB2, and SMAD3 were observed. Traditionally, miRNAs function to silence or inhibit their direct gene targets; therefore, these inverse correlations were as predicted and further establish that, by inhibiting miRNAs in circulation, functionality of target genes might be altered at the transcriptional level, suggesting potential relevance for future countermeasure development. The cross-species, cross-tissue comparison was motivated by translational considerations: while the liver is a central organ for space-related metabolic and fibrotic risk, peripheral blood is the most practical biospecimen for longitudinal monitoring in astronauts. Our goal was therefore not to equate mouse liver and human blood responses, but to determine whether similar TGF-β and miRNA-associated signatures emerge in the readily accessible human compartment.
Fig. 9
Spearman correlation plots showing correlations between 14 studied miRNAs and 11 mRNAs using data from the NASA Twin Study. Plots generated for various white blood cell types, including PBMCs A, LD B, CD19 C, and CD4 D
Comments (0)