
Remember me

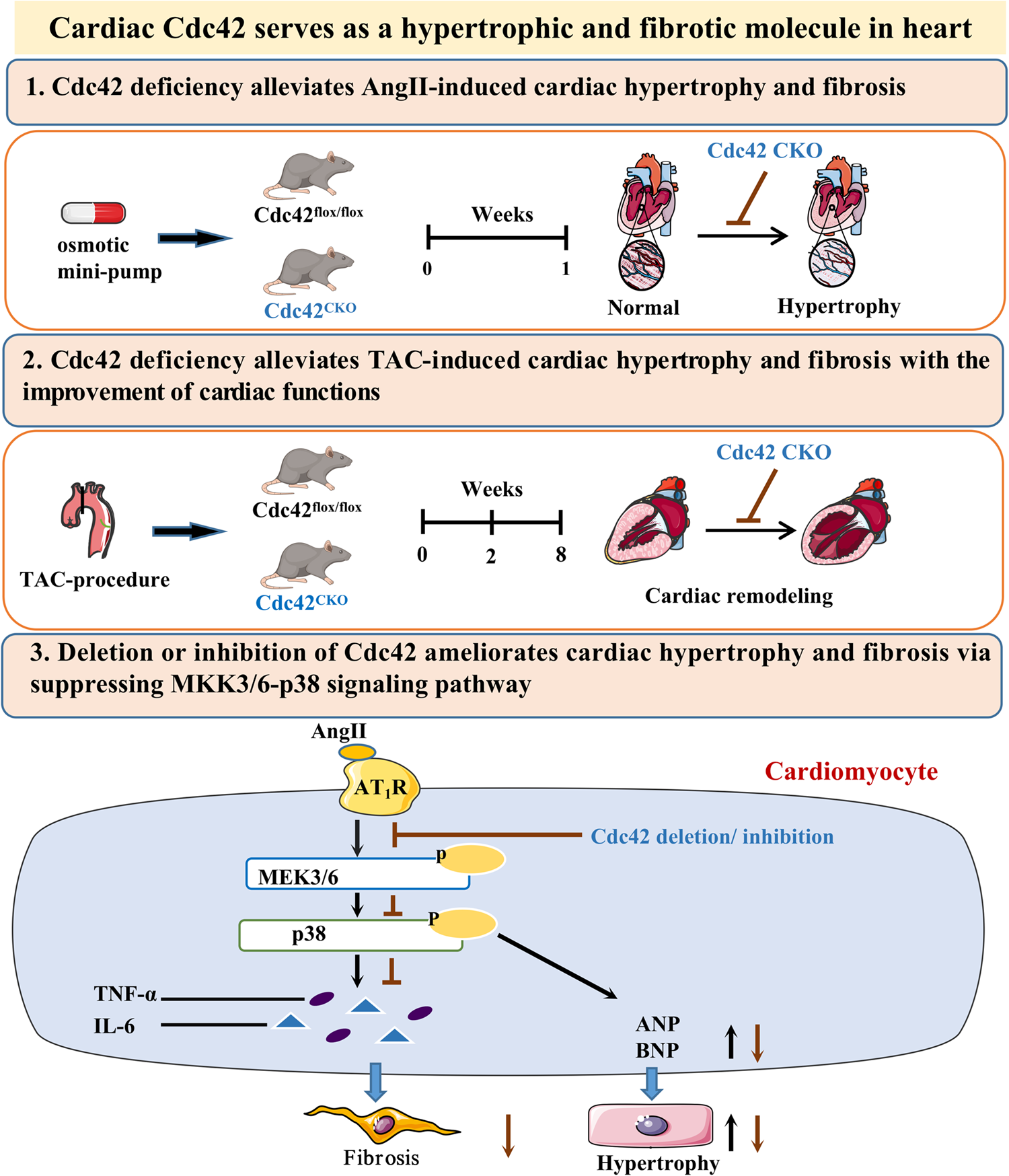
Glycans are not primary gene products but secondary products of a complex interplay among glycogenes. The epigenetic and expression regulation of glycogenes provides a deeper understanding of glycomic complexity. In previous studies, primary ESCC tumor tissues and matched adjacent nonneoplastic tissues from 155 ESCC patients were subjected to WGS, WGBS, RNA-seq and proteomics analysis [23]. Using the above multi-omics data, we performed a multi-omics analysis to study the global variation profiles of 641 glycogenes downloaded from the UniProt database.
Transcriptome analysis revealed significant differences in the expression of 276 glycogenes (fold change > 1.5 and P < 0.05) in tumors compared with paired normal samples, among which 173 and 103 genes were upregulated and downregulated, respectively (Supplementary Table 3). Integration analysis of the epigenome and transcriptome was applied to screen for glycogenes whose differential expression was caused by promoter DNA methylation in ESCC. The stringent calling criteria (average |Δβ|≥ 0.15, fold change > 1.5 and P < 0.05; Benjamini–Hochberg corrected P < 0.05, detailed in Methods) and manual selection led to the identification of three glycogenes exhibiting strong and consistent methylation-driven transcriptional upregulation or downregulation (Supplementary Fig. 1). For example, ALG1L is associated with a decrease in promoter hypermethylation and greater ALG1L gene expression in ESCC tissues than in matched normal tissues (Supplementary Fig. 1A–C). The relationships between gene expression and ALG1L methylation were directly inversely correlated (Supplementary Fig. 1D). Conversely, epigenetic downregulation due to significant promoter hypermethylation was observed in ADAMTS5 and GALNT5 (Supplementary Fig. 1E–L).
Due to differences in the lymph node status of 155 ESCC samples (82 N0, 41 N1, 22 N3, and 10 N4 samples), we existed interested in glycogen dynamics during ESCC progression, particularly lymph node metastasis (LNM). We performed dynamic cluster analysis basis of glycogene quantity to explore the types of dynamic changes. We acquired an array of glycogene clusters whose fold change increased or decreased as cancer developed with lymph node metastasis.
A total of 13 clusters were detected in 155 ESCC patients, and four glycogen clusters showed an obvious expected change trend (Supplementary Fig. 1M). Cluster #7 and cluster #11 represented an array of glycogenes whose expression quantity presented an increasing trend with the progression of the disease, whereas cluster #2 and cluster #9 represented glycogene clusters with a decreasing trend in expression quantity (Supplementary Fig. 1M). KEGG-based enrichment analysis revealed that glycogenes involved in mannose type O-glycan biosynthesis and glycosphingolipid biosynthesis-globo and isoglobo series tended to increase, while other types of O-glycan biosynthesis and glycosaminoglycan biosynthesis-keratan sulfate tended to decrease in the process of lymph node metastasis (Supplementary Fig. 1N). In addition, the glycogenes in clusters #7 and #11 were mostly involved in base excision repair, sphingolipid metabolism, other glycan degradation, and lysosomes (Supplementary Fig. 1N). There are relatively few glycogenes in the “quantity decreased” clusters. In addition, significant biological processes involved in both increased and decreased glycogene clusters were also identified, including glycosphingolipid biosynthesis-lacto and neolacto series, mucin type O-glycan biosynthesis, glycosaminoglycan degradation, fructose and mannose metabolism, and so on (Supplementary Fig. 1N). Details of the glycogene clusters and affected glycogenes are shown in Supplementary Table 4.
Copy number dysregulation-mediated alterations of glycogenes in ESCCIntegration analysis of the transcriptome and WGS was applied to identify glycogenes whose differential expression was associated with copy number variation and somatic mutation. Overall, none of the differentially expressed glycogenes were significantly or frequently mutated. Therefore, we explored the relationships between expression levels and CNVs in all 155 individuals across all the glycogenes. The default thresholds of + 0.2 and −0.25 log2 ratios were used to detect single-copy gains or losses in ESCC tissues compared with matched normal tissues. Thirty glycogenes were selected for correlation analysis, and the expression levels of seven glycogenes were positively correlated with estimates of copy numbers (Spearman’s rank correlation rho > 0.3; P < 0.05). In this analysis, the ALG3, B4GALT4, DPY19L4, EDEM2, POFUT1, RPN1 and XXYLT1 glycogenes were identified (Fig. 1A, B).
Fig. 1
Correlation in copy number and expression levels across seven glycogenes. A Heatmap of DNA copy number and gene expression. The upper panel represents the DNA copy number. The bottom represents the RNA expression. B RNA expression values and DNA copy number values are plotted against each other for the 155 cases where WGS and RNA sequencing were performed
Then we analyzed the expression changes of these genes (ALG3, B4GALT4, DPY19L4, EDEM2, POFUT1, RPN1, XXYLT1) in ESCC across multi-omics data (including our previously published transcriptome data [23], Liu's published proteome data [24], and our undergoing proteome data). Our analysis showed that in Liu's published proteome data, the expression of ALG3 (log2FC = 1.045682), DPY19L4 (log2FC = 0.51101), EDEM2 (log2FC = 0.17026) in ESCC was significantly higher than that in normal tissues. However, the log2FC values of DPY19L4 and EDEM2 were both lower than 0.585, indicating that the differential expression multiple of DPY19L4 and EDEM2 in ESCC and normal tissues was less than 1.5. Meanwhile, the expression of ALG3, DPY19L4 and EDEM2 is not detected in our unpublished proteome data. In our undergoing proteome data, the expression of XXYLT1 (log2FC = 0.085533) in ESCC was significantly higher than that in normal tissues, but the log2FC values of it was lower than 0.585, indicating that the differential expression of XXYLT1 in ESCC and normal tissues was less than 1.5. The expression of B4GALT4 in ESCC and normal tissues were inconsistent in the three data sets (Supplementary Fig. 2C–E). The expression of POFUT1 and RPN in ESCC was higher than that in normal tissues in all three data sets (Figs. 2A, 6A). Therefore, in the end, we chose POFUT1 and RPN1 with more significant differences (P < 2.22e − 16) for follow-up research.
Fig. 2
POFUT1 is highly expressed and is a poor prognostic factor in ESCC. A POFUT1 expression in ESCC and normal tissues from Liu’s published proteome, our unpublished proteome data and previously published transcriptome data. B Representative images and quantification of POFUT1 expression in the cytoplasm and nucleus of ESCC tissues and paired normal tissues (n = 85) detected by IHC. Scale bar, 100 μm. C Kaplan–Meier plot presenting the correlation between high POFUT1 expression and poor prognosis in ESCC patients and in male ESCC patients, patients with a history of alcohol consumption, and patients with stage T3. D Univariate and multifactorial Cox regression analyses of the association of POFUT1 levels with clinicopathological factors in ESCC patients
POFUT1 encodes an O-fucosyltransferase that specifically modifies epidermal growth factor-like domains (EGF-LD) and transfers fucose to threonine or serine residues within the O-fucosylation motif [25]. The protein encoded by RPN1 is part of the N-oligosaccharyl transferase complex, which binds high mannose oligosaccharides to asparagine residues in the Asn-X-Ser/Thr consensus motif of nascent polypeptide chains [26].
POFUT1 is highly expressed and a poor prognostic factor in ESCCWe first investigated POFUT1 expression via multi-omics data. Our results showed that the expression of POFUT1 in ESCC was obviously higher than that in normal tissues according to Liu’s published proteome [24] and our unpublished proteome data, as well as previously published transcriptome data [23] (P < 0.001) (Fig. 2A). POFUT1 protein expression in ESCC was further assessed by Microarray-based immunohistochemistry. Our results suggested that POFUT1 was expressed mainly in the cytoplasm of ESCC, samples and that its expression was significantly higher in the cytoplasm of ESCC samples than in paired normal esophageal squamous epithelium samples (P < 0.001) (Fig. 2B). ESCC nuclear expression was not significantly different from that of normal tissues (Fig. 2B).
Patients were categorized into a high POFUT1 expression group (≥ 98.9655) and a low POFUT1 expression group (< 98.9655) by applying median POFUT1 expression in the cytoplasm. The results of the correlation analysis between POFUT1 expression and clinicopathologic characteristics are was shown in Supplementary Table 5. Survival analysis demonstrated that the mean survival time was 942 days for patients with high POFUT1 expression and 1 224 days for patients with low POFUT1 expression. Patients with high POFUT1 expression had a significantly shorter survival time than did those with low POFUT1 expression (P < 0.05) (Fig. 2C). This trend was also observed in male patients, patients with a history of alcohol consumption, and patients with stage T3 (P < 0.05) (Fig. 2C). Univariate and multifactorial Cox regression analyses revealed that high POFUT1 expression was an important risk factor for poor prognosis in ESCC patients (Fig. 2D).
Knockdown of POFUT1 inhibits the proliferation, metastasis and invasion of ESCC.To further investigate the role of POFUT1 in ESCC, we first investigated the endogenous expression of POFUT1 in 6 ESCC cell lines. As shown in Fig. 3A, POFUT1 was expressed at a lower level in KYSE450 cells and at a higher level in KYSE150, TE-1, TE-5, TE-9, and TE-14 cells. KYSE150 and TE-1 cells with high endogenous POFUT1 expression were used for POFUT1 knockdown, and the knockdown efficiency was validated by RT-qPCR and Western Blot. The results showed that the mRNA and protein expression of POFUT1 were significantly lower in KYSE150 and TE-1 cells transfected with siRNAs than in the negative control cells (Fig. 3B, C). MTT, colony formation, and transwell assays showed that POFUT1 knockdown significantly inhibited the proliferation, colony formation, migration and invasion of KYSE150 and TE-1 cells (Fig. 3D–F).
Fig. 3
Knockdown of POFUT1 suppresses the proliferation, metastasis and invasion of ESCC. A POFUT1 protein expression was examined in 6 kinds of cells by Western Blot. B, C RT-qPCR (B) and Western Blot (C) revealed that POFUT1 was efficiently knocked down in KYSE150 and TE-1 cells. D, E Analysis of the proliferation ability of POFUT1 knockdown KYSE150 and TE-1 cells by MTT (D) and colony formation assays (E). F Analysis of the migration and invasion abilities of POFUT1-knockdown KYSE150 and TE-1 cells by transwell assays. G Analysis of E-cadherin, N-cadherin and vimentin expression in POFUT1-knockdown KYSE150 and TE-1 cells by Western Blot. Scale bar, 200 μm. **, P < 0.01; ***, P < 0.001
Epithelial-mesenchymal transition (EMT) is a biological process that involves in epithelial cells acquiring mesenchymal features, during which cells exhibit loss of epithelial markers (E-cadherin, ZO-1) and gain of mesenchymal markers (N-cadherin, vimentin) [27]. EMT closely relates to tumor invasion and metastasis [28]. To determine whether the EMT process was affected by POFUT1, we detected the protein expression of EMT markers in ESCC cells with POFUT1 knockdown or overexpression. The results showed that POFUT1 had no effect on E-cadherin expression in KYSE150 and TE-1 cells (Fig. 3G). However, the knockdown of POFUT1 in KYSE150 and TE-1 cells reduced N-cadherin and vimentin expression (Fig. 3G).
To further confirm the effect of POFUT1 on ESCC proliferation, we performed a subcutaneous tumor formation experiment in nude mice using the KYSE150 cell line. The knockdown efficiency was validated by RT-qPCR and Western Blot. The results showed that the mRNA and protein expression of POFUT1 were significantly lower in KYSE150 cells transfected with shRNA than in the negative control cells (Supplementary Fig. 3A, B). The tumors of the mice in the stably POFUT1 knockdown group (POFUT1-sh) grew more slowly and had a smaller tumor volume than did those in the empty vector group (nc group) (Supplementary Fig. 3C–E). These results suggested that the knockdown of POFUT1 inhibited the proliferation, metastasis and invasion of ESCC in vivo and in vitro.
POFUT1 overexpression promotes the proliferation, metastasis and invasion of ESCC.Subsequently, we overexpressed POFUT1 in KYSE450, KYSE150 and TE-1 cells, and the overexpression efficiency was verified by RT-qPCR and Western Blot (Fig. 4A, B). The overexpression of POFUT1 enhanced the proliferation and colony formation of KYSE450 cells (Fig. 4C, D). The overexpression of POFUT1 significantly enhanced the migration and invasion of KYSE450, KYSE150 and TE-1 cells (Fig. 4E). We also detected the protein expression of EMT markers in POFUT1-overexpression ESCC cells. The results showed that POFUT1 had no effect on E-cadherin expression in KYSE450 and TE-1 cells (Fig. 4F). However, POFUT1 overexpression in KYSE450 and TE-1 cells increased N-cadherin and vimentin expression (Fig. 4F).
Fig. 4
POFUT1 overexpression promotes the proliferation, metastasis and invasion of ESCC. A, B RT-qPCR (A) and Western Blot (B) revealed that POFUT1 was efficiently overexpressed in KYSE450, TE-1 and KYSE150 cells. C, D Analysis of the proliferation of POFUT1-overexpression KYSE450 cells by MTT (C) and colony formation assays (D). E Analysis of the migration and invasion abilities of POFUT1-overexpression KYSE450, TE-1 and KYSE150 cells by transwell assays. F Analysis of E-cadherin, N-cadherin and vimentin expression in POFUT1-overexpression KYSE450 and TE-1 cells by Western Blot. G Quantitation of the number of lung metastatic nodules in mice in the NC group and POFUT1-OE group. H Representative images and HE staining of lung tissues from mice in the NC group and POFUT1-OE group. Scale bar, 200 μm. *, P < 0.05; **, P < 0.01; ***, P < 0.001
To further confirm the effect of POFUT1 on ESCC invasion and migration, we constructed a tail vein metastasis model using the KYSE150 cell line. Compared with those in the empty vector group (NC group), the mice of the stably overexpressing POFUT1 group (POFUT1-OE) exhibited significantly increased lung metastatic nodules, which was also confirmed by the quantitative analysis and HE staining (P < 0.05) (Fig. 4G, H). These results suggested that the overexpression of POFUT1 promoted the metastasis of ESCC in vitro and in vivo.
Overexpression of POFUT1 increases overall fucosylation levels and activates the Notch signaling pathway, which mediates POFUT1 induced pro-migration in ESCCPOFUT1 is primarily responsible for adding fucose residues to specific glycoproteins through O-linkage. To reveal the underlying mechanism of POFUT1 in ESCC, we measured the total level of fucosylation by click chemistry reaction. Fucosylated proteins were labeled with Click-iT® fucose alkyne and then the biotin-azide was attached to the fucosylated proteins by a click chemistry reaction. The results visualized by the HRP-streptavidin antibody showed that the level of overall cellular fucosylation was significantly reduced after POFUT1 knockdown in KYSE150 and TE-1 cells (Fig. 5A). Conversely, the overexpression of POFUT1 significantly increased the overall level of fucosylation in KYSE450 and TE-1 cells (Fig. 5B).
Fig. 5
The overexpression of POFUT1 increases the overall O-fucosylation level and activates the Notch signaling pathway, which mediates POFUT1 induced pro-migration in ESCC. A Analysis of the level of overall cellular O-fucosylation modification after POFUT1 knockdown in KYSE150 and TE-1 cells by click chemistry reaction. B Analysis of the level of overall O-fucosylation modification after POFUT1 overexpression in KYSE450 and TE-1 cells by click chemistry reaction. C Analysis of the protein level and fucosylation modification level of Notch1 in KYSE450 and TE-1 cells stably overexpressing POFUT1 and the control using AAL lectin enrichment followed by Western Blot. D, E Analysis of the protein expression of NICD (D) and HES1 (E) after POFUT1 was knocked down in KYSE150 and TE-1 cells, and POFUT1 was overexpressed in KYSE450 and TE-1 cells. F Analysis of the migration and invasion abilities of KYSE450 and TE-1 cells stably overexpressing POFUT1, with or without DAPT treatment by transwell assay. **, P < 0.01; ***, P < 0.001
Notch1 is a transmembrane receptor containing an extracellular domain with 36 tandem EGF repeats [29]. Considering the specific modification effect of POFUT1 on EGF domains, we selected Notch1 as a downstream target of POFUT1 for mechanistic studies in ESCC. The protein levels and fucosylation modification levels of Notch1 in POFUT1-overexpressing and control cells were measured by AAL lectin enrichment followed by Western Blot. The results indicated that POFUT1 overexpression significantly increased the fucosylated levels of Notch1 in KYSE450 and TE-1 cells, while the protein expression levels increased to a lesser extent than the fucosylation modification levels did (Fig. 5C). Grayscale analysis also confirmed that the fold change of the increase in Notch1 fucosylation was significantly higher than that in Notch1 protein expression (Fig. 5C). Considering that Notch1 is a core member of the Notch pathway, which plays an important role in modulating the cellular malignant phenotype, we aimed to explore whether POFUT1 modulated the Notch signaling pathway in ESCC. Interestingly, POFUT1 knockdown in KYSE150 and TE-1 cells markedly decreased the protein expression of NICD and its downstream target gene HES1 (Fig. 5D, E). POFUT1 overexpression in KYSE450 and TE-1 cells enhanced protein expression of NICD and HES1 (Fig. 5D, E). Collectively, our results suggest that POFUT1 regulates the activation of the Notch signaling pathway in ESCC.
We further confirmed that POFUT1 promotes the migration and invasion of ESCC via the Notch signaling pathway. We added DAPT, a Notch inhibitor, to POFUT1-overexpressing ESCC cells to observe whether the addition of DAPT could reverse the promoting effect of elevated POFUT1 expression on migration and invasion of ESCC cells. First, we verified the inhibitory effect of 5μMol DAPT on the Notch signaling pathway by Western blot and its influence on the proliferation of ESCC cells by MTT assay. Our results suggest that 5μMol DAPT can decrease the expression of Notch1, NICD and HES1 and inhibit the proliferation of KYSE450 and TE-1 cells (Supplementary Fig. 4A, B). The results indicated that the addition of DAPT to POFUT1 overexpressing ESCC cells reversed the promoting effect of elevated POFUT1 expression on the migration and invasion of ESCC cells (Fig. 5F). Therefore, the Notch signaling pathway may partially mediate the oncogenic role of POFUT1 in ESCC.
RPN1 glycogene alterations and potential oncogenic effects in ESCC.In addition to POFUT1, RPN1 is a glycogene that shows significant copy number gain driven transcriptional and protein upregulation. RPN1 was completely covered by the significantly amplified focal region at approximately 8q24.3 and harbored significant amplification in ESCC with WGS. Its copy number amplification was significantly correlated with RNA expression (Spearman’s correlation coefficient > 0.3; P < 0.05) (Fig. 1B). Furthermore, the mRNA and protein expression of RPN1 in ESCC was upregulated compared with that in paired normal tissues according to transcriptomic, published and unpublished proteomic data [23, 24] (Fig. 6A). Western Blot analysis consistently confirmed differences in RPN1 expression between ESCC and paired normal tissues (Fig. 6B). The immunohistochemistry results from the microarrays show that RPN1 protein expression in the cytoplasm and nucleus of ESCC cells was significantly higher than that in non-tumor tissues (Fig. 6C). However, there was no obvious correlation in ESCC patients according to different sex, age, clinical stage, tumor size, etc. (Supplementary Table 6). Kaplan–Meier analysis indicated that ESCC patients with high RPN1 expression had a worse prognosis than did those with low RPN1 gene expression (P < 0.05) (Fig. 6D). This trend was also observed in male patients, patients with grades 1–2, and patients with stage T3 (P < 0.05) (Fig. 6D).
Fig. 6
RPN1 is highly expressed and is a poor prognostic factor in ESCC. A RPN1 expression in ESCC and normal tissues from Liu’s published proteome, our unpublished proteome data and previously published transcriptome data. B The protein expression of RPN1 was analyzed by Western Blot in ESCC and paired normal tissues. C Representative images and staining quantification analysis of RPN1 expression in the cytoplasm and nucleus of ESCC tissues and paired normal tissues (n = 85) detected by IHC. Scale bar, 100 μm. D Kaplan–Meier plot presenting the correlation between high RPN1 expression and poor prognosis in ESCC patients and male ESCC patients, patients with G1-2, and patients with stage T3
To investigate the biological role of RPN1 in ESCC, we detected the endogenous expression of RPN1 in seven ESCC cell lines and found that RPN1 was expressed at a lower level in TE-5 cells than in KYSE150, KYSE450, TE-1, and TE-14 cells (Fig. 7A). KYSE150 and KYSE450 cells with high RPN1 expression were transfected with the siRNA targeting RPN1 and the negative control plasmid. As shown in Fig. 7B, the knockdown efficiency was verified by Western Blot in KYSE150 and KYSE450 cells. MTT, colony formation, and transwell assay showed that RPN1 knockdown significantly inhibited the proliferation, colony formation, invasion and migration of KYSE150 and KYSE450 cells (Fig. 7C, D, F). In addition, RPN1 knockdown increased cell apoptosis in KYSE150 and KYSE450 cells by flow cytometry (Fig. 7E).
Fig. 7
RPN1 knockdown suppresses proliferation, metastasis and invasion, and promotes apoptosis in ESCC cells. A RPN1 protein expression was examined in 5 kinds of cells by Western Blot. B Western Blot revealed that RPN1 was efficiently knocked down in KYSE150 and KYSE450 cells by Western Blot. C, D Analysis of the ability of proliferation in RPN1 knockdown KYSE150 and KYSE450 cells by MTT (C) and colony formation assays (D). E Analysis of the apoptosis in RPN1 knockdown KYSE450 and KYSE150 cells by flow cytometry. UR (upper right) refers to the upper right of the four regions divided by cross coordinates in the flow diagram. This area represents the percentage of late apoptotic cells. LR (lower right) refers to the lower right of the four regions divided by cross coordinates in the flow diagram. This area represents the percentage of early apoptotic cells. % Gated (UR + LR) refers to the percentage of early and late apoptotic cells. F Analysis of the ability of migration and invasion in RPN1 knockdown KYSE450 and KYSE150 cells by transwell assay. **P < 0.01; ***, P < 0.001. G Bubble diagram of KEGG-based enrichment analysis was performed for those 137 genes whose expression significantly differed
It has been reported that the ribophorin protein is involved in multiple signaling pathways involved and the biological processes in cancer. To examine the signaling pathway and detailed mechanism of RPN1 in ESCC, we performed RNA sequencing in the RPN1 knockdown and negative control groups of KYSE150 cells. Three independent biological replicates were conducted. The results showed that 71 genes were upregulated and 66 genes were downregulated according to the transcriptome data with a fold change > 1.5 and P < 0.05 (Supplementary Table 7). KEGG-based enrichment analysis was performed for those 137 genes whose expression significantly differed and revealed that the TNF signaling pathway, p53 signaling pathway, ErbB signaling pathway, and so on were altered significantly (Fig. 7G). These results suggest that RPN1 regulation in the malignant progression of ESCC may be related to the TNF signaling pathway, p53 signaling pathway, ErbB signaling pathway, etc.
Comments (0)