
Remember me

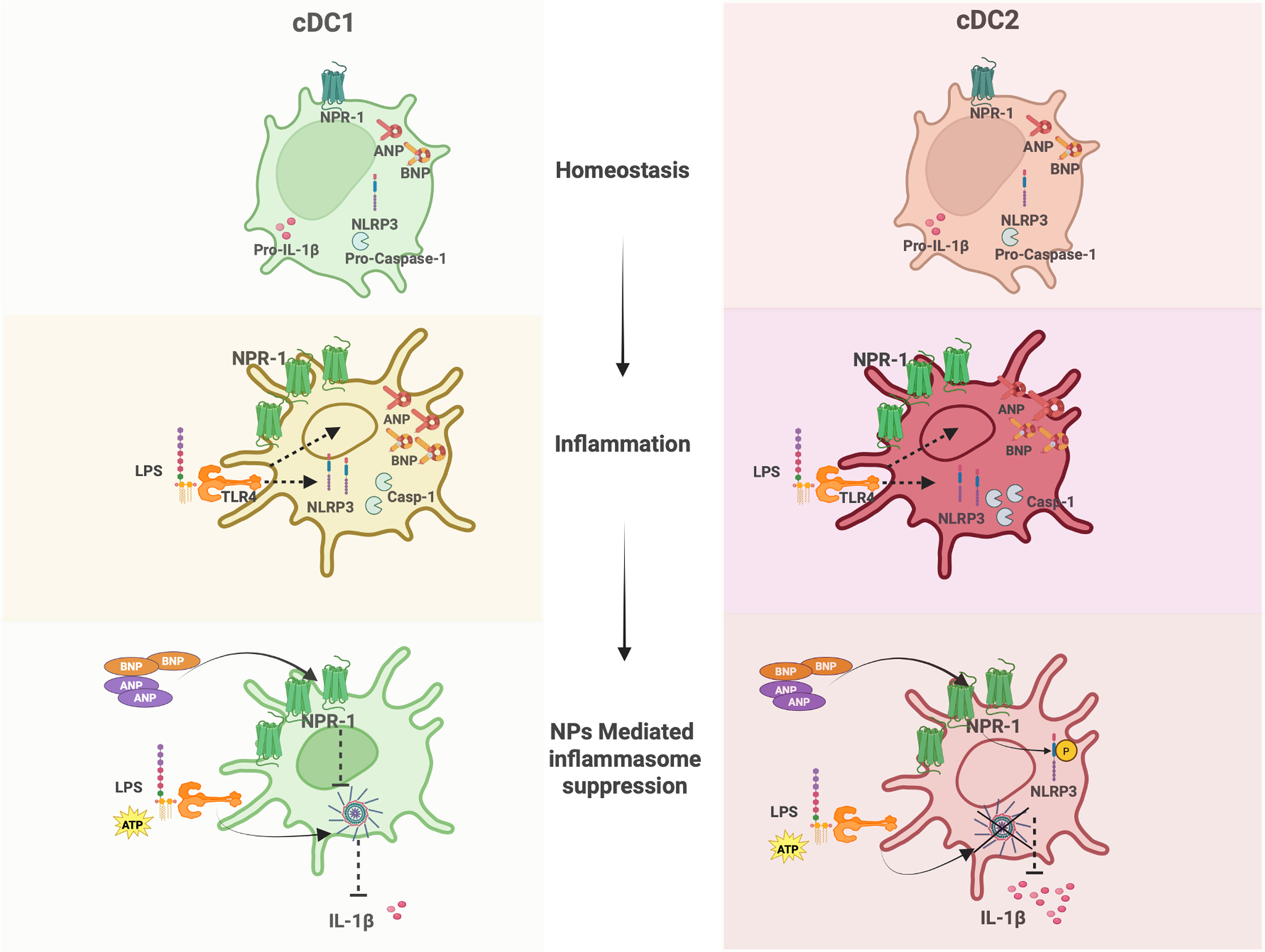
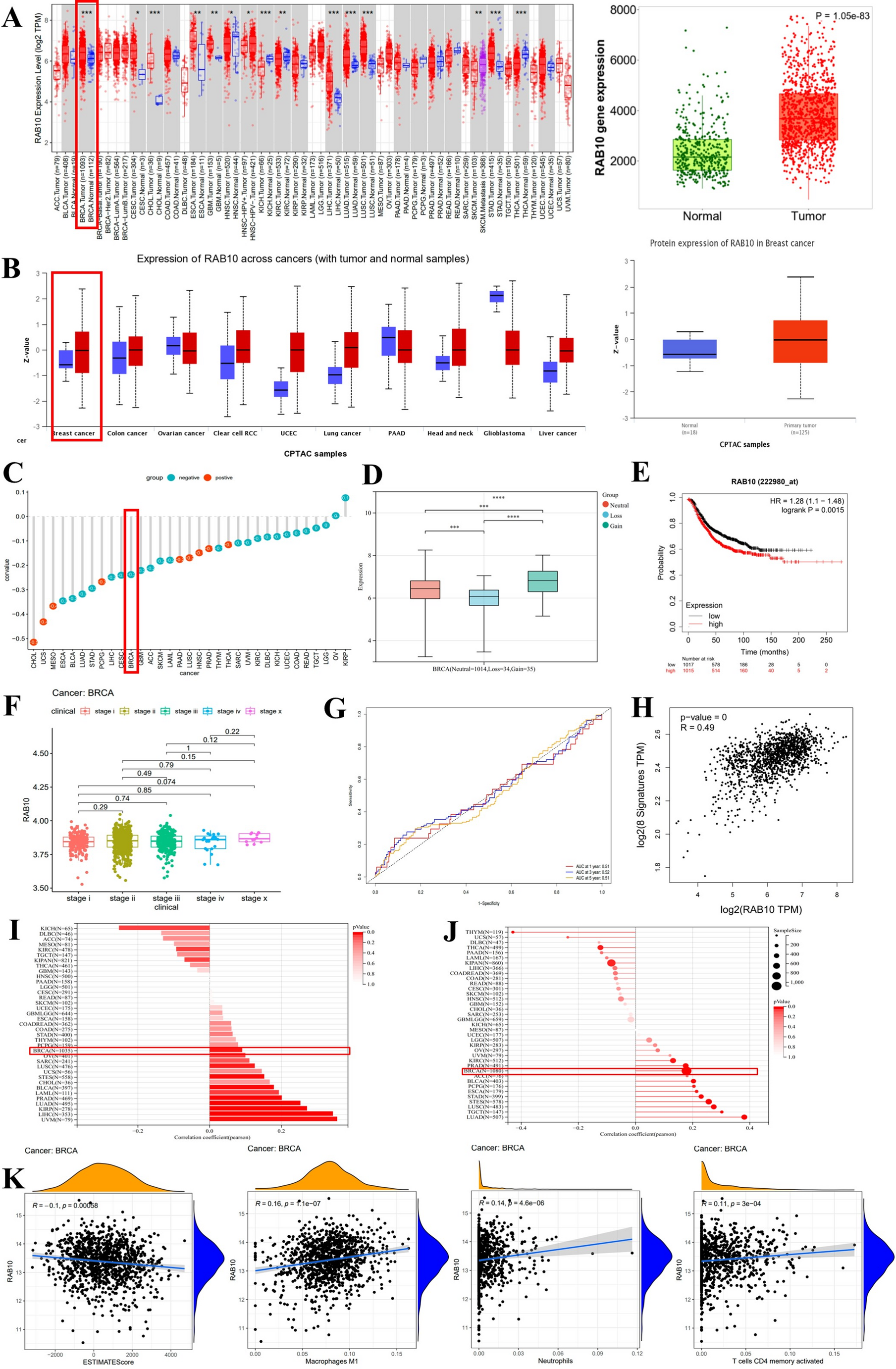
A previous study demonstrated that human pDCs express NPR1 [28], highlighting the potential involvement of the NP/NPR1 axis in immune regulation. To investigate the role of ANP and BNP in modulating NLRP3 inflammasome activation in cDCs, we examined the NPs/NPR1 expression profile in murine cDC1 and cDC2 subsets. Microarray data (GSE203450) [30], were re-analyzed for genes critical to the NPs axis, including Npr1, Nppa, and Nppb. No significant differences were observed in the mRNA expression levels of Nppa, Nppb, or the receptor gene Npr1 in purified bone marrow-derived cDC1 and cDC2 under steady-state conditions, as illustrated by the Volcano plot (Fig. 1a). Key genes of the NPs/NPR1 axis from the same dataset were further analysed and showed as heatmaps comparing cDC1 and cDC2 both at steady state (control) and in inflammatory conditions (i.e., following 24 h LPS treatment). The data showed that LPS stimulation induced Npr1 mRNA expression more prominently in cDC2 than in cDC1, while Nppa and Nppb expression remained relatively unchanged (Fig. 1b).
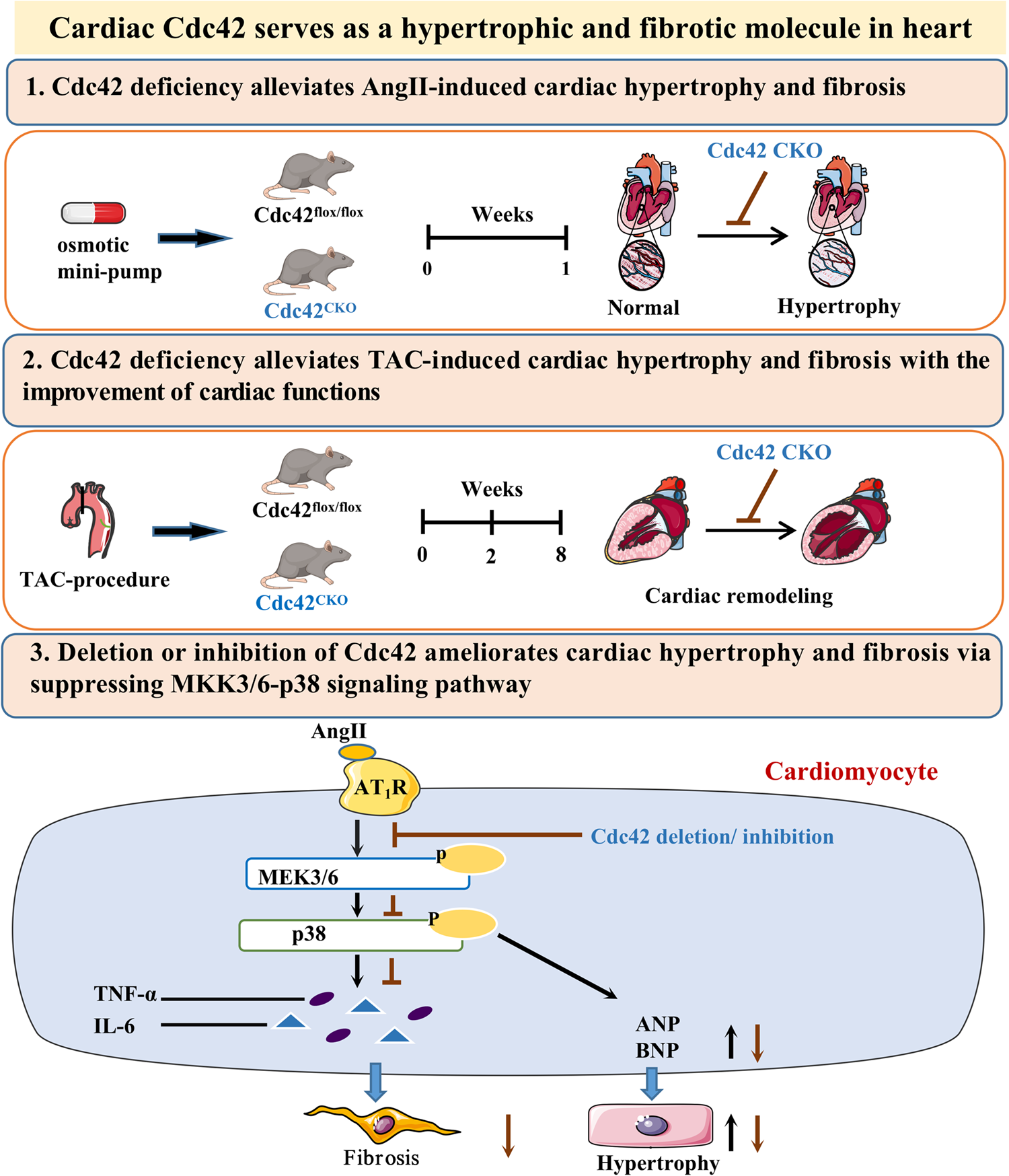
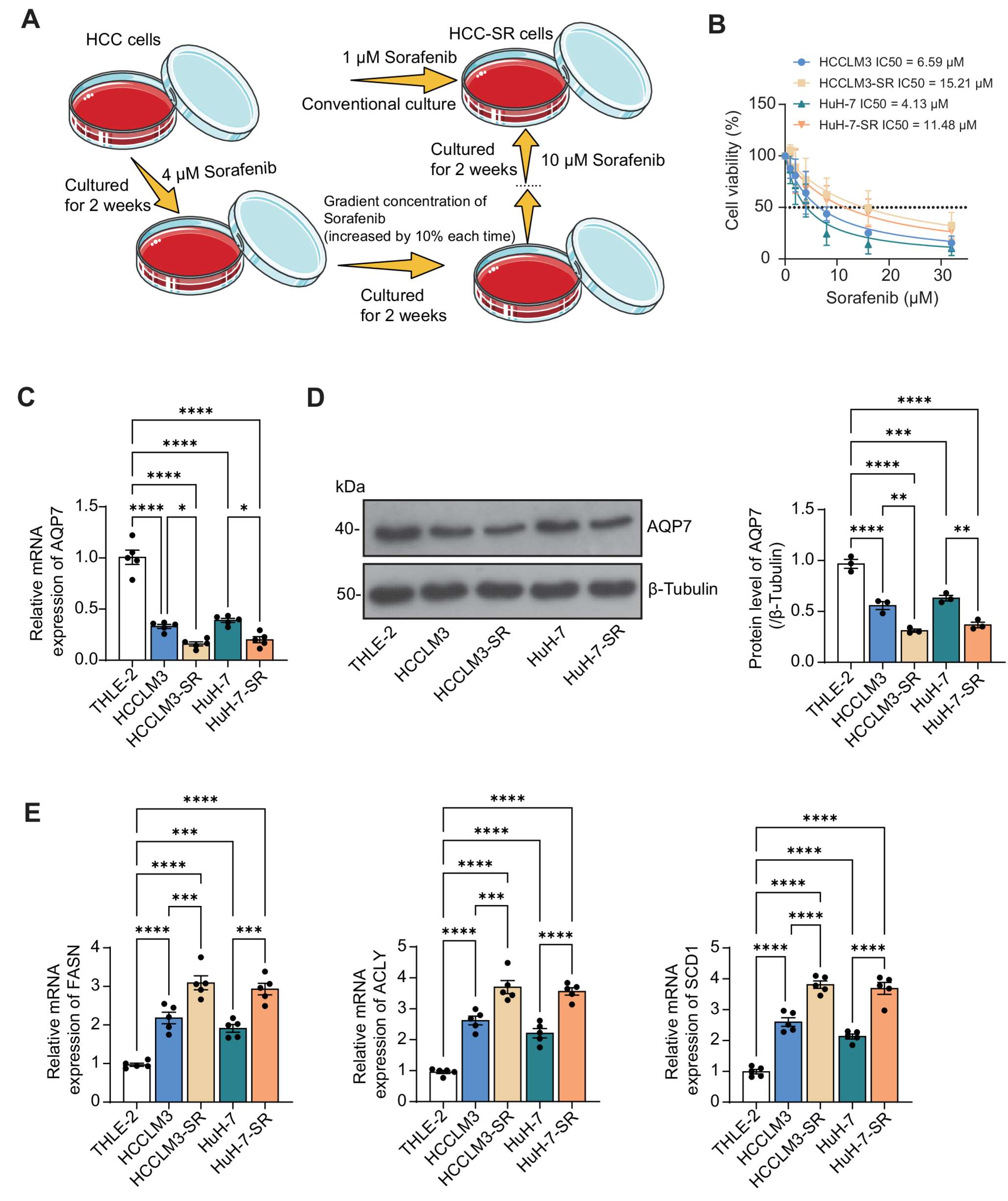
Fig. 1
Natriuretic Peptide Axis profile expression in cDC subsets. a The volcano plot shows differentially expressed genes between steady-state cDC1 and cDC2 derived from bone marrow cells, highlighted by colors (enriched in cDC1, blue; enriched in cDC2, red; not significantly enriched, gray). b The heatmap shows the expression of z-scores of genes related to NPs axis in bone marrow derived cDC1 and cDC2 under steady-state condition and after stimulation with LPS for 24 h. c, d Mutu cDC1 and cDC2 cells were cultured for 24 h. Medium was replaced, and cells were treated with LPS (100ng/ml for 24 h). ANP, BNP and NPR1 gene c and protein d expression were assessed by quantitative RT-PCR and WB analysis, respectively. The values were normalized to β-tubulin. Data are represented as mean ± SD, with n = 3. Untreated cells were used as control and assumed as 1; p < 0.05 vs untreated cells by unpaired t-test
We next assessed Npr1, Nppa, and Nppb gene and protein expression in mutu cDC1 and cDC2 cell lines. Real-time PCR confirmed that both mutu cDC1 and cDC2 expressed the Npr1 receptor, Nppa, and Nppb transcripts under steady-state conditions, with no significant changes in response to LPS stimulation (Fig. 1c). At the protein level, both mutu cDC1 and cDC2 were found to express the NPR1 receptor, and a slight increase in its expression was observed after 24 h LPS stimulation particularly in cDC2. Notably, ANP levels showed a marked increase upon LPS stimulation in both types of cDCs, while BNP protein levels exhibited a modest increase upon LPS stimulation in both cDCs (Fig. 1d).
These results demonstrate that cDC1 and cDC2 are sources of both ANP and BNP under inflammatory conditions, accompanied by a slight increase in NPR1 receptor expression. This suggests that these endogenous peptides may regulate cDCs functions through both endocrine, but also autocrine loops.
cDC1 and cDC2 cells display differential inflammasome activationPrevious studies have demonstrated that NPs can modulate inflammasome activation in myeloid cells, particularly in human monocytes [5]. However, inflammasome expression and activation in DCs remains uncharacterized. To address this, we re-analyzed, as shown in Fig. 1a, the murine dataset of bone marrow derived cDCs [30], to assess the expression of inflammasome-related genes in the two main cDC subsets under steady-state conditions and following LPS stimulation.
Analysis of inflammasome-related genes, visualized using a volcano plot, revealed their expression in both cDC1 and cDC2 subsets (Fig. 2a). Notably, under steady-state conditions, mRNA transcripts encoding sensor proteins (e.g., Aim2, Nlrp1a, Nlrp3, Pycard, and others) were more abundant in cDC1, whereas those encoding effector proteins (e.g., Casp1, Casp4, Il1b, and Gsdmd) were enriched in cDC2 (Fig. 2a, b). To further assess how these transcripts respond to inflammatory conditions, we analyzed key inflammasome-related genes from the same dataset and visualized their expression as heatmaps. We found that LPS treatment induced expression of effector proteins in cDC2, but not in cDC1, and not significant transcriptional induction in sensor proteins in both cDCs subsets. Transcripts encoding sensors proteins appeared to be slightly downregulated in cDC1 upon 24 h LPS stimulation (Fig. 2b).
Fig. 2
Differential NLRP3-inflammasome activation in cDC subsets. a The volcano plot shows differentially expressed steady-state genes between cDC1 and cDC2 derived from bone marrow cultures. Key inflammasome genes differentially expressed were highlighted by colors (enriched in cDC1, blue; enriched in cDC2, red; not significantly enriched, gray). b The heatmap shows the expression of z-scores for genes related to inflammasome in bone marrow derived cDC1 and cDC2 under steady-state conditions and after stimulation with LPS for 24 h. c Mutu cDC1 and cDC2 cells were cultured for 24 h. Medium was replaced, and cells were treated with LPS (100ng/ml for 24 h). Nlrp3, Caspase-1 and Il-1β gene expression were assessed by quantitative RT-PCR. d Schematic representation of inflammasome activation and analysis in cDC subsets (Created by BioRender.com). cDC1 and cDC2 cells were grown for 24 h. Medium was replaced, and cells were stimulated with LPS (100ng/ml for 60 min) and ATP (5 mM for 30 min) (i.e.LPS + ATP) e–f. NLRP3 and Caspase-1 protein expression levels in cDC1 e and cDC2 f were assessed by WB and normalized to β-tubulin. IL-1β protein secretion was measured by ELISA in culture supernatants in cDC1 e and cDC2 f. Data are represented as mean ± SD, with n = 3;*p < 0.05 vs untreated cells by unpaired t-test; Data are represented as mean ± SD, with n = 3. Untreated cells were used as control and assumed as 1; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001 vs untreated cells by unpaired t-test or by two-way ANOVA
To further validate these findings, mutu cDCs were either left untreated or stimulated with LPS for 24 h, followed by analysis of Nlrp3, Casp1, and Il1b expression using real-time PCR. At steady state, mutu cDC1 and cDC2 exhibited comparable levels of these inflammasome-related genes. However, LPS stimulation for 24 h led to a significant increase in Nlrp3 and Il1b expression, predominantly in the mutu cDC2 subset (Fig. 2c). Next, mutu cDC1 and cDC2 were stimulated with LPS and ATP (LPS + ATP), a classical trigger of NLRP3 inflammasome activation in macrophages, and inflammasome activation was assessed as outlined in Fig. 2d. LPS + ATP treatment resulted in differential activation of the inflammasome pathway between the two cDC subsets. Specifically, cDC2 cells exhibited robust inflammasome activation, as indicated by elevated protein levels of the NLRP3 receptor and the active p20 fragment of Caspase-1, compared to cDC1 cells. In line with these results, LPS + ATP stimulation led to higher IL-1β secretion in cDC2 cells compared to cDC1 cells (Fig. 2e, f).
Collectively, these findings indicate that the cDC2 subset is highly equipped to respond to inflammatory stimuli and exhibits a markedly stronger NLRP3 inflammasome activation upon LPS + ATP treatment compared to cDC1.
NPs/NPR1 axis repress inflammasome activation in cDCsBased on previous findings [5, 6, 31] and the observed expression of inflammasome-related genes and proteins in cDCs (Fig. 2a, b), we hypothesized that exogenous NPs might influence LPS + ATP-induced inflammasome activation in these cells. To test this, cDC1 and cDC2 cells were stimulated in vitro, and inflammasome activation was assessed as shown in Fig. 3a. Notably, brief exposure of cDCs to ANP or BNP (e.g. 10 min prior to LPS + ATP stimulation) effectively inhibited early Caspase-1 activation and significantly reduced IL-1β secretion at 24 h in both cDC1 and cDC2 subsets (Fig. 3b, c). This effect was more pronounced in cDC2 cells compared to cDC1, as expected (Fig. 3b, c).
Fig. 3
NPs/NPR1 axis repress NLRP3-Inflammasome activation in cDCs. a Graphical representation of the experimental model (Created by BioRender.com). cDC1 and cDC2 cells were grown for 24 h, medium was replaced, and cells were treated with ANP and BNP (0.1μM for 10 min) and then stimulated with LPS (100ng/ml for 60 min) and ATP (5 mM for 30 min) (LPS + ATP) and Caspase-1 protein expression was assessed by WB analysis b and the level of IL-1β in culture supernatants was determined by ELISA c. cDC1 and cDC2 cells were pre-treated with H-3048 (10μM for 10 min) before (LPS + ATP) stimulation and Caspase-1 activation d or IL-1β secretion e in culture supernatants were assessed by WB or ELISA, respectively. Untreated cells were used as control and assumed as 1, n = 3; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001 vs untreated cells; #p < 0.05; # # # #p < 0.0001; LPS + ATP + ANP or BNP vs H-3048 + LPS + ATP + ANP or BNP treated cells; by one-way ANOVA followed by Tukey’s multiple comparisons test
To determine whether the NPs/NPR1 axis played a role in this inhibitory effect, we pretreated cDCs with H-3048, a potent NPR1 antagonist, before ANP or BNP exposure and subsequent LPS + ATP stimulation. H-3048 strongly reversed the NPs-mediated inhibition of LPS + ATP-induced Caspase-1 activation in both subsets (Fig. 3d). Interestingly, in cDC2 cells, H-3048 pretreatment also increased constitutive Caspase-1 activation (Fig. 3d). Regarding IL-1β secretion, H-3048 fully abrogated the inhibitory effect of NPs in cDC2 cells (Fig. 3e).
Overall, these findings demonstrate that the NPs/NPR1 axis suppresses inflammasome activation in cDCs, with a more pronounced inhibitory effect in cDC2 cells.
NPs/NPR1 axis repress priming and oligomerization of NLRP3-Inflammasome platform in cDCsCanonical NLRP3 activation is a tightly regulated process that occurs in two distinct steps. The first step, considered rate-limiting, involves the upregulation of NLRP3 protein levels [32], while the second step includes inflammasome oligomerization, driven by post-translational modifications including NLRP3 phosphorylation [20, 21]. We found that both ANP and BNP reduced LPS + ATP-induced NLRP3 expression in cDC2 cells, an effect that was completely reversed by H-3048 (Fig. 4a). A similar trend was observed in cDC1 cells, although the changes were not statistically significant (Fig. 4a).
Fig. 4
NPs/NPR1 axis represses priming and oligomerization of NLRP3-Inflammasome platform in cDCs. cDC1 and cDC2 cells were grown for 24 h. Medium was replaced, and cells were treated with ANP and BNP (0.1μM for 10 min) and then stimulated with LPS (100ng/ml for 60 min) and ATP (5 mM for 30 min) (LPS + ATP) in absence or presence of H-3048 pretreatment (10μM for 10 min). NLRP3 a and phospho-NLRP3 (Ser295) (p-NLRP3) b protein expression was assessed by WB analysis. For NLRP3 analysis untreated cells were used as control and assumed as 1; for p-NLRP3 analysis LPS + ATP cells were used as control and assumed as 1, n = 3; *p < 0.05; ***p < 0.001 versus untreated or LPS + ATP stimulated cells by one-way ANOVA followed by Tukey’s multiple comparisons test
To further investigate whether NPs modulate NLRP3 activation through post-translational mechanisms, we assessed the effect of ANP and BNP on NLRP3 phosphorylation levels in LPS + ATP-treated cDC1 and cDC2 cells. Notably, we found that pre-treatment with ANP and BNP enhanced NLRP3 phosphorylation (Fig. 4b).
These findings indicate that the NPs/NPR1 axis negatively impact on both the priming and oligomerization steps of inflammasome activation in cDCs, highlighting its role as a new key checkpoint pathway, particularly in the cDC2 subset.
Comments (0)