
Remember me
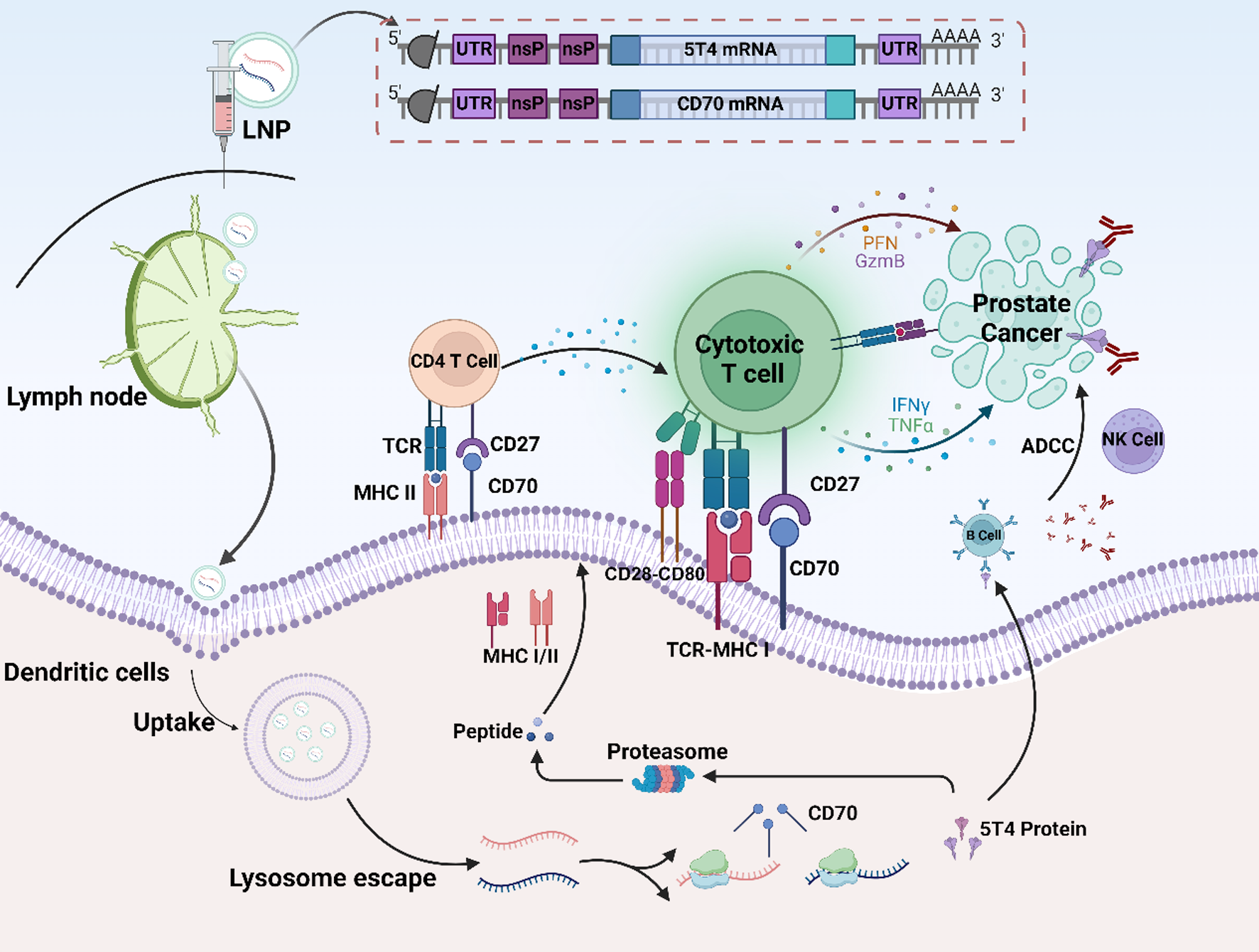

Initially, the open reading frame (ORF) sequences of the target genes 5T4 and CD70 were optimized, and the corresponding mRNAs were synthesized in vitro (Fig. 1A). The synthesized mRNAs were subsequently transfected into bone marrow-derived dendritic cells (BMDCs), and western blotting and flow cytometry analyses confirmed the successful expression of 5T4 and CD70, thereby validating the efficient transfection and expression of the target genes (Fig. 1B-C). Next, the target mRNA was encapsulated in RNA-lipid nanoparticles (RNA-LNPs) using E12A1A3, an innovative ionizable lipid that has been previously developed and characterized [33]. Dynamic light scattering (DLS) analysis of the resulting LNPs revealed that the particle diameter ranged from 90 to 100 nm, with a polydispersity index (PDI) of less than 0.2, indicating a uniform size distribution (Fig. 1D). Additionally, the zeta potential of the LNPs stabilized at approximately + 5 mV, with the mRNA encapsulation efficiency exceeding 90% (Figure S1A-B). Further experiments confirmed that the mRNA encapsulated within the LNPs maintained excellent stability during storage, with minimal degradation and retained functionality over time (Figure S1C-D; Figure S2A-C). After validating the physicochemical properties of the LNPs, the biosafety of mRNA-LNPs was assessed. BMDCs were exposed to various concentrations of mRNA-LNPs in a concentration-dependent manner. The results indicated that BMDCs exhibited optimal growth conditions during co-transfection, with a maximum tolerable concentration of 15 µg/ml (Fig. 1E). Therefore, 1 µg/ml was chosen for subsequent experiments to prevent significant apoptosis of BMDCs (Fig. 1F-G). Safety assessments were performed across all treatment groups to ensure that mice demonstrated good tolerance to therapy. Blood biochemical analyses were performed to monitor potential systemic toxicity. The results showed that Key markers of liver and kidney function, including alanine aminotransferase (ALT), aspartate aminotransferase (AST), blood urea nitrogen (BUN), and creatinine (CREAT), remained within normal physiological ranges across all groups, indicating no signs of hepatic or renal dysfunction (Fig. 1H). These findings suggested that the administration of LNP-based formulations, including CD70-LNPs and 5T4-LNPs, was well tolerated without inducing measurable biochemical toxicity. To further corroborate these observations, histological examinations of the major organs (the heart, liver, kidneys, spleen, and lungs) were conducted. Hematoxylin and eosin (H&E) staining of tissue sections revealed no significant histopathological changes or tissue damage in any treatment group (Figure S3) After confirming both in vitro and in vivo safety of 5T4-LNPs, their immunogenicity was further assessed. In healthy C57BL/6 mice, three immunizations were performed, and T cells were isolated from the spleen and co-cultured with BMDCs pretreated with various formulations. IFN-γ production was measured using ELISpot assay (Fig. 1I). The results demonstrated that BMDCs pretreated with 5T4-LNPs had significantly enhanced IFN-γ production compared to control BMDCs, indicating that 5T4-LNPs effectively activated specific immune responses, highlighting their potential as tumor-associated antigen mRNA vaccines (Fig. 1J-K). This study demonstrated the successful optimization, synthesis, and delivery of 5T4 and CD70 mRNA via LNPs, confirming their efficient expression in dendritic cells, in vitro and in vivo safety, and ability to activate immune responses, highlighting their potential as tumor-associated antigen mRNA vaccines.
Fig. 1
Preparation, characterization and immunogenicity of mRNA-LNP. (A) Schematic structure of 5T4 and CD70 mRNAs; (B-C) The protein expression of 5T4 and CD70 mRNAs was verified by western blot and flow cytometry, respectively; (D) Hydrodynamic sizes of 5T4 and CD70 LNPs measured by dynamic light scattering; (E) Assessing the cytotoxic effects of increasing concentrations of mRNA-LNPs on BMDCs using the CCK-8 assay; (F-G) The percentage of apoptotic cells induced by 5T4/CD70-LNPs at 1 µg/ml was observed in the right quadrant using the Annexin V-PE/7-AAD apoptosis kit; (H) ALT, AST, BUN, and CREAT blood testing results after the mRNA-LNPs immunization (I) IFN-γ spot-forming cells (J) and statistical data (K) from restimulated splenocytes determined by the ELISPOT assay on day 10 after 5T4-LNPs immunization. Data indicate mean ± SEM of biological replicates (n ≥ 3 per group).Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
Maturation and activation of BMDCs and cytokine levels in T cells after in vitro treatment with different combinations of 5T4 and CD70 mRNA-LNPsAfter completing the safety validation of 5T4-LNP and CD70-LNP, and assessing the immunogenicity of 5T4-LNP, we further investigated the phenotypic modulation of BMDCs by these LNPs. Phenotypic changes in BMDCs were evaluated 24 h after transfection with 5T4-LNPs, CD70-LNPs, or their combination. The results showed that both 5T4-LNPs and CD70-LNPs significantly increased the expression of the proliferation marker Ki-67 (Fig. 2A), indicating enhanced cell proliferation. Additionally, both treatments upregulated the levels of the antigen-presenting molecule MHC-II, suggesting improved antigen-presenting capacity of the transfected BMDCs. Moreover, CD70-LNPs notably upregulated key co-stimulatory molecules, such as CD70, CD40, and the maturation markers CD80 and CD86 (Fig. 2B-F). When combined with 5T4-LNPs, the upregulation of these molecules was even more pronounced, suggesting a synergistic effect on the activation and maturation of dendritic cells. Further analysis revealed an increase in the expression of functional regulatory markers CD103, CD69, and OX40L, which enhanced the immunogenic characteristics of the transfected BMDCs (Fig. 2G-I). Subsequently, after 48 h of co-culture between T cells and BMDCs treated with different LNP formulations, only the combination group elicited robust secretion of TNF-α and IFN-γ by T cells, indicating that dual treatment is essential for fully activating the cytotoxic capacity of T cells (Fig. 2J-M). However, the expression of the T cell regulatory molecules 4-1BBL and B7-H2, as well as the DC migration marker CCR7, did not change significantly (Figure S4). These results suggested that although transfection promoted the activation and maturation of BMDCs, its impact on their migratory potential or specific regulatory pathways was not significant. These findings indicate that 5T4-LNPs and CD70-LNPs effectively enhanced the proliferation, antigen-presenting capacity, and co-stimulatory molecule expression of BMDCs, with a synergistic effect observed when both formulations were combined, indicating their potential to enhance dendritic cell activation and maturation for improved immunotherapeutic efficacy. Building upon these results, future studies will investigate the functional implications of these phenotypic changes in eliciting robust antitumor immune responses.
Fig. 2
Maturation and activation of BMDCs and cytokine levels in T cells after in vitro treatment with different combinations of 5T4 and CD70 mRNA-LNPs. BMDCs transfected with 1 µg/ml mRNA-LNPs were stained after 24 h for the proliferation-related marker Ki-67 (A), antigen presentation function, and costimulatory molecules CD70, MHC-II, CD40, CD86 (B-F), CD80, and DC maturation and functional regulation markers CD103, CD69, and OX40L (G-I). Expression levels of TNF-α and IFN-γ in T cells (J-M). The percentage of positive cells is indicated, and each symbol represents the BMDCs from one mouse. Data are presented as mean ± SEM of biological replicates (n ≥ 3 per group). Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
CD70 significantly enhanced the humoral and cellular immunity initiated by 5T4-LNPs in vivomRNA vaccines are recognized for their ability to induce robust immune responses through activation of both humoral and cellular immunity. Based on the phenotypic modulation of BMDCs by 5T4-LNPs and CD70-LNPs, the roles of these two mRNA-LNPs in in vivo immune response were investigated. To evaluate the specific role of CD70-LNPs in driving these immune responses, immunization experiments were conducted in tumor-free C57BL/6 mice. After three rounds of immunization, blood samples were collected for serum analysis and T cells were isolated from the spleen for functional assessment (Fig. 3A). Serum analysis revealed a significant increase in 5T4-specific antibodies in mice treated with CD70-LNPs, suggesting that CD70-LNPs play a key role in enhancing humoral immune response (Fig. 3B). To further assess the cellular immune response, the activation status of CD8 + T cells was analyzed. Compared to the group treated with only 5T4-LNPs, mice treated with a combination of 5T4-LNPs and CD70-LNPs showed a 15–20% increase in the secretion of IFN-γ and TNF-α from CD8 + T cells, indicating a synergistic effect of the combined treatment in enhancing the activation and function of cytotoxic T cells (Fig. 3C-F). Additionally, RM-1 cells and their transplanted tumors expressed the 5T4 protein (Figure S5A). To further evaluate the cytotoxic potential of these activated CD8 + T cells, they were co-cultured with the prostate cancer cell line RM-1. The combined treatment group exhibited the highest cytotoxic efficiency, significantly enhancing the killing of RM-1 target cells compared to the 5T4-LNPs-only group (Fig. 3G-H). However, after the second stimulation of BMDCs with 5T4-LNPs, no significant difference in splenic T cell proliferation was observed between the combined treatment group and the 5T4-LNPs-only group (Figure S5B). This enhanced cytotoxicity underscores the potential of CD70-LNPs to boost the antitumor activity of cytotoxic T lymphocytes, further demonstrating the dual role of CD70-LNPs in promoting both humoral and cellular immunity.
Fig. 3
CD70 significantly enhanced the humoral and cellular immunity initiated by 5T4-LNPs in vivo. (A) The timeline and experimental groups for 5T4 and CD70 mRNA-LNPs administration in tumor-free male mice; (B) Relative levels of 5T4-specific IgG in sera of immunized mice on day 10 post-immunization; Representative flow cytometry analysis and mean percentage of IFN-γ (C, D) and TNF-α expression (E, F) in CD8+ T cells from the spleens of mice post-immunization; (G-H) Representative flow cytometry analysis and killing efficiency of T cells from the spleens of mice post-immunization. Data indicate mean ± SEM of biological replicates (n ≥ 3 per group).Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
CD70 mRNA-LNPs facilitates anti-tumor efficacy in the B16-OVA and pancreatic cancer modelsBased on the results presented above, the potential of CD70 as a target for tumor immunotherapy will be further validated and its role in promoting antitumor immune responses will be assessed. The B16-OVA model is a widely used murine model for evaluating antitumor immune responses and vaccine efficacy, particularly in the study of tumor immunotherapies and vaccines. This model provides an experimental platform for assessing immune system responses to OVA by combining the B16 melanoma cell line with recombinant ovalbumin (OVA) [34, 35]. This study elucidated the mechanisms by which CD70-LNPs enhance immune responses in a B16-OVA classical tumor model. OVA-specific antibody titers were measured regularly throughout the treatment (Fig. 4A-B). The results showed that The total IgG levels in the combined treatment group (OVA + CD70-LNPs) were consistently higher than those in the OVA-LNP group, suggesting that CD70-LNPs play a pivotal role in enhancing the humoral immune response. Notably, no significant changes in body weight were observed in any treatment group, suggesting that all treatments were well tolerated with no systemic toxicity observed. Both OVA-LNPs and OVA + CD70-LNPs treatments significantly inhibited tumor growth, with the combined treatment group showing notably better results than the single-treatment groups (Fig. 4C-E). The tumor volume in the combined treatment group remained consistently below 500 mm³, with a continuous decline throughout the study, further demonstrating the enhanced antitumor activity of CD70-LNPs (Figure S6A-D). Immune cell infiltration analysis of the tumor microenvironment revealed a slight increase in CD8 + T cell infiltration in the OVA + CD70-LNPs group compared to that in the OVA-LNPs or CD70-LNPs groups (Fig. 4F-G). Although no significant change was observed in the proportion of tetramer-positive T cells (Fig. 4H), the NK cell activation markers CD69 and CD107a were significantly upregulated in the OVA + CD70-LNPs group (Fig. 4I-J). This upregulation was strongly correlated with the observed increase in IgG levels, suggesting that the combined treatment effectively enhanced NK cell activation. Additionally, an increased proportion of NK cells and CD69-positive cells in the spleen was observed, along with an increase in CD4 + and CD8 + T cell populations (Figure S7A-D). To further investigate the mechanism by which CD70-LNPs enhance specific IgG production, a detailed analysis of splenic immune response was conducted. The results showed that in mice treated with CD70-LNPs, there was a significant increase in the number of germinal center B cells (B220 + CD95 + GL-7+), plasma cells (B220 + CD138+) (Fig. 4K-N), and helper T cells (Figure S7E-F). These findings suggest that CD70 plays a key role in enhancing antibody-dependent cellular cytotoxicity (ADCC) through NK cell activation, significantly promoting the overall cellular immune response. Consequently, these enhanced immune responses translate into stronger tumor suppression and sustained immune activation. Furthermore, CLDN18.2 is considered an effective target for pancreatic cancer treatment, has been validated in CAR-T, ADC, and mRNA vaccines [36, 37]. To further assess the therapeutic effect of CD70-LNPs in non-classical models, a subcutaneous Panc02 tumor mouse model was established. The results showed that the combination of CLDN18.2-LNPs and CD70-LNPs yielded the most significant tumor suppression, significantly prolonging the survival of mice, with no noticeable adverse effects in any of the treatment groups (Fig. 4O-Q, Figure S8A-D). These observations suggested that CD70 plays a pivotal role in enhancing ADCC, likely through NK cell activation, and significantly contributes to the overall cellular immune response. Consequently, these enhancements translate into improved tumor suppression and robust immune activation.
Fig. 4
CD70 mRNA-LNPs facilitates anti-tumor efficacy in the B16-OVA and pancreatic cancer models. (A) The timeline for OVA and CD70 mRNA-LNPs administration in B16-OVA tumor-bearing female mice; (B) Relative levels of OVA-specific IgG in the serum of immunized mice on days 6, 10, and 14 during the course of immunization; (C) Body weight variation curves of mice during the immunization period; (D) Average tumor growth curves of B16-OVA under different treatment groups; (E) The tumors of each group were photographed 19 days after tumor inoculation; (F-G) Flow cytometry analysis and percentages of tumor-infiltrating CD8+ T cells in each group following different treatments; (H) Representative flow dot plots and statistical data of the H-2Kb/SIINFEKL tetramer staining of CD3+CD8+ T cells in the tumor post-immunization; (I-J) Representative flow cytometry plots and statistical data of CD69 and CD107a staining in NK cells from tumors post-treatment; (K-L) Flow cytometry analysis of spleens post-immunotherapy showing the proportion of germinal center B cells (GL-7+ CD95+); (M-N) Representative flow cytometry plots and statistical data of mature B cells in the spleen post-treatment. (O-Q) Average tumor growth curves of Panc02, survival curves for mice and body weight variation curves of mice under different treatment Data indicate mean ± SEM of biological replicates (n ≥ 3 per group).Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
CD70 mRNA-LNPs enhances the immunotherapeutic efficacy of the 5T4 mRNA-LNPs in the prostate cancer mouse modelThe immunogenicity of 5T4 and its specific cytotoxicity against RM-1 cells in vitro have been confirmed, and CD70 has demonstrated potent immune-enhancing efficacy in various tumor models. Next, we evaluated the therapeutic efficacy of a combination of 5T4 and CD70-LNPs in a mouse model of prostate cancer. Following administration of the five LNPs, the combination group exhibited a significant advantage in both tumor volume reduction and overall survival. Tumor volumes consistently remained below 500 mm³ in the combination group mice, demonstrating superior antitumor ability compared to the other groups. Notably, compared to the control group, the combined therapy significantly increased the average survival time, indicating substantial therapeutic benefits. (Fig. 5A-H). Given the enhanced ADCC and cellular immune responses previously observed with CD70-LNPs, we further analyzed the effects of combination therapy on T and NK cell populations in both the spleen and tumor-infiltrating immune cells. The combination group showed a significant increase in the proportion of CD4 + and CD8 + T cells in the spleen, indicating systemic immune activation (Fig. 5I-O). However, the most pronounced increase was observed in tumor-infiltrating CD8 + T cells and NK cells, suggesting that the therapy effectively boosted effector cell infiltration into the tumor microenvironment (Fig. 5P-S). The combination group exhibited the highest levels of IFN-γ and TNF-α secretion by tumor-infiltrating CD8⁺ T cells (Fig. 5T-U). While no significant changes were noted in tumor-infiltrating dendritic cells, a slight reduction in splenic DCs was observed, possibly reflecting specific immune modulation (Fig. S9A-C). Immunofluorescence staining further supported these findings, revealing elevated expression of the effector cytokines IFN-γ and TNF-α at the tumor site in the combination group. This indicates that CD70 plays a dual role in enhancing both humoral and cellular immunity, contributing to the observed tumor suppression (Fig. S9D-E). To better understand the broader immune remodeling induced by the therapy, we analyzed splenic immune cell populations. CD8 + T cells from treated mice exhibited significantly increased expression of the stem cell marker TCF-7 and effector molecules granzyme B and perforin, indicating enhanced cellular immunity (Fig. 6A-C). Similarly, TCF-7 expression was significantly elevated in CD4 + T cells without a corresponding increase in regulatory T cell (Treg) population (Fig. S10A-B), suggesting that therapy did not promote an immunosuppressive state. This finding was further supported by the stable expression of CTLA-4 (Fig. S10C-D), indicating that CD70-LNPs selectively enhanced effector T-cell responses without driving immunosuppressive pathways. Moreover, combination therapy increased the proportion of central memory T cells (TCM) in the spleen, implying that CD70-LNPs promote the generation of long-lasting memory CD8 + T cells, which are crucial for sustained antitumor immunity (Fig. 6D-G). Beyond T cell responses, splenic DCs exhibited increased expression of the maturation markers CD86 and MHC-II, which is consistent with the activation of BMDCs observed in vitro, further supporting the conclusion that CD70-LNPs not only activated T cells but also enhanced antigen presentation by both macrophages and DCs (Fig. 6H-K). Additionally, CD70-LNPs significantly influenced macrophage polarization. M1 macrophages displayed elevated MHC-II expression, indicating an enhanced antigen presentation (Fig. 6L-M). The therapy promoted macrophage differentiation toward the pro-inflammatory M1 phenotype (F4/80+/CD206-) (Fig. 6N-P), as confirmed by immunofluorescence staining of tumor sections (Fig. 6Q). In conclusion, CD70-LNPs not only facilitate TAA-specific immunity, but also enhance macrophage inflammatory polarization to inhibit tumor progression, and the alliance of these effects results in the anti-tumor efficiency of CD70 and TAA-LNPs.
Fig. 5
CD70 mRNA-LNPs enhances the immunotherapeutic efficacy of the 5T4 mRNA-LNPs in the prostate cancer mouse model. (A) The timeline of 5T4 and CD70 mRNA-LNPs administration in RM-1 tumor-bearing male mice; (B-C) Tumor growth curves for RM-1 tumors and survival curves for mice under different treatment groups; (D) The tumors of each group were photographed 19 days after tumor inoculation; (E-F) Individual tumor growth curves within each treatment group; (I-L) Representative dimensionality reduction plots of NK cells (green), CD4+ T cells (blue) and CD8+ T cells (red) in the spleen under different treatment groups; (M-O) Statistical data on the proportion of NK cells, CD4+ T cells and CD8+ T cells in the spleen post-treatment; (P-Q) Flow cytometry analysis and percentages of tumor-infiltrating CD8+ T cells in each group following different treatments; (R-S) Flow cytometry analysis and percentages of tumor-infiltrating NK cells in each group following different treatments; (T-U)Flow cytometry analysis of the percentage of TNF-α and IFN-γ expressing CD8⁺ tumor-infiltrating T cells following different treatments. Data are presented as mean ± SEM of biological replicates (n ≥ 3 per group). Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
Fig. 6
CD70 promotes positive regulation of the overall immune system in a prostate cancer mouse model. Representative flow cytometry plots and statistical data of TCF-7 (A), Granzyme B (B), and perforin (C) expression in CD8+ T cells in the spleen under different treatment groups; representative flow cytometry plots and statistical data of TCM in CD4+ (D-E) and CD8+ T cells (F-G) in the spleen under different treatment groups; (H-I) representative flow dot plots and statistical data of MHC II staining of macrophage cells in the spleen under different treatment groups; representative flow cytometry plots and statistical data of antigen presentation function and costimulatory molecules CD86 (J-K) and MHC-II (L-M) expression in splenic dendritic cells; (N-P) representative flow cytometry plots and statistical data of macrophage phenotypes (anti-inflammatory mφ and pro-inflammatory mφ) in the spleen under different treatment groups; (Q) Confocal fluorescence images of tumor tissues in the following treatment groups. The nuclei, F4/80, and CD206 were stained blue (DAPI), yellow, and purple, respectively. Data are presented as mean ± SEM of biological replicates (n ≥ 3 per group). Error bars represent 95% CI. ****P < 0.0001; ***P < 0.001; **P < 0.01; *P < 0.05; ns., not significant
Bulk RNA sequencing results of dendritic cells in the spleen and T cells in the draining lymph nodes under different treatment groupsBulk RNA sequencing (RNA-seq) was performed on DCs isolated from the spleen and T cells isolated from the draining lymph nodes of mice administered with LNPs (Fig. 7A). Differential gene expression analysis of splenic DCs revealed a marked upregulation of genes associated with key DC subsets. Specifically, genes linked to conventional DC1 (cDC1) differentiation, such as Tlr3, Cadm1, Irf8, Strip2, and Clec9a, were elevated. These genes are essential for antigen cross-presentation and priming of cytotoxic T lymphocytes (CTLs), emphasizing the critical role of cDC1 cells in antitumor immunity. Plasmacytoid DCs (pDCs) are known for their ability to produce large quantities of type I interferons, which contribute to both antiviral and antitumor responses. Genes related to pDCs, including Spib, Ly6c2, Runx2, Siglech, and Cd209a, were significantly upregulated in the 5T4 + CD70 group, indicating an enhanced pDC differentiation. Furthermore, genes associated with the conventional DC2 (cDC2) lineage, including Cd22, Cd4, Usp18, and Lyz1, were upregulated. Given the crucial role of cDC2 in promoting the development of helper T cells and B cells, these genes may play a pivotal role in polarizing Th2 responses and facilitating B cell development, thereby mediating (ADCC) (Fig. 7B-D). These findings highlight that CD70-5T4 LNPs modulate DC differentiation, particularly favoring cDC1 and pDC lineage development, both instrumental in initiating and sustaining CTL responses that are vital for effective antitumor immunity. In parallel, RNA-seq analysis of T cells isolated from draining lymph nodes of the combination group mice revealed a significant increase in transcriptomic signatures associated with memory CD8 + T cells as well as Th1 and Th17 T cell populations, which are key drivers of cellular immunity (Fig. 7E). The upregulation of transcriptional regulators involved in T cell receptor (TCR) signaling, including Nfkb1, Nfatc2, and Fos, reflects robust activation of TCR-mediated pathways critical for T cell proliferation, activation, and differentiation into effector populations capable of anti-tumor activity. Moreover, increased expression of genes related to T-cell differentiation and function, such as Runx3, Cd44, and Bcl6, has been observed, promoting effector and memory T-cell development. Concurrently, effector molecules, including Ifng (encoding interferon gamma), Prf1 (encoding perforin), and Gzmb (encoding granzyme B), were upregulated, underscoring the role of T cell-mediated cytotoxicity in tumor clearance. Upregulation of co-stimulatory molecules and chemokine receptors, such as Cd27 and Cxcr6, further indicated enhanced T cell activation, migration, and interaction within the tumor microenvironment (Fig. 7F-J). These results collectively demonstrate that 5T4-CD70 combination therapy promotes DC differentiation toward lineages essential for initiating CTL responses while simultaneously driving robust T cell activation and differentiation into effector cells. This dual activation of both DC and T cell compartments underscores the potent immunostimulatory effects of combination therapy, contributing to enhanced antitumor immunity.
Fig. 7
Bulk RNA sequencing results of dendritic cells in the spleen and T cells in the draining lymph nodes under different treatment groups. (A) The timeline of 5T4 and CD70 mRNA-LNPs administration in RM-1 tumor-bearing male mice and the immune cell isolation strategy; Heatmap presenting the altered expression of cDC1-associated (B), cDC2-associated (C), and pDC-associated (D) genes in dendritic cells isolated from the spleens of different treatment groups; (E) Proportions of T cell type clusters in draining lymph nodes across different treatment groups as identified by RNA-sequencing based on CIBERSORT; (F-J) Expression levels of selected gene modules (TCR signaling, Activation/differentiation, Effector molecules, Co-stimulatory receptors and Chemokine receptors) in T cells from draining lymph nodes after treatment with different therapies, as determined by RNA sequencing. Log2 values of the FPKM values were plotted
Comments (0)