
Remember me
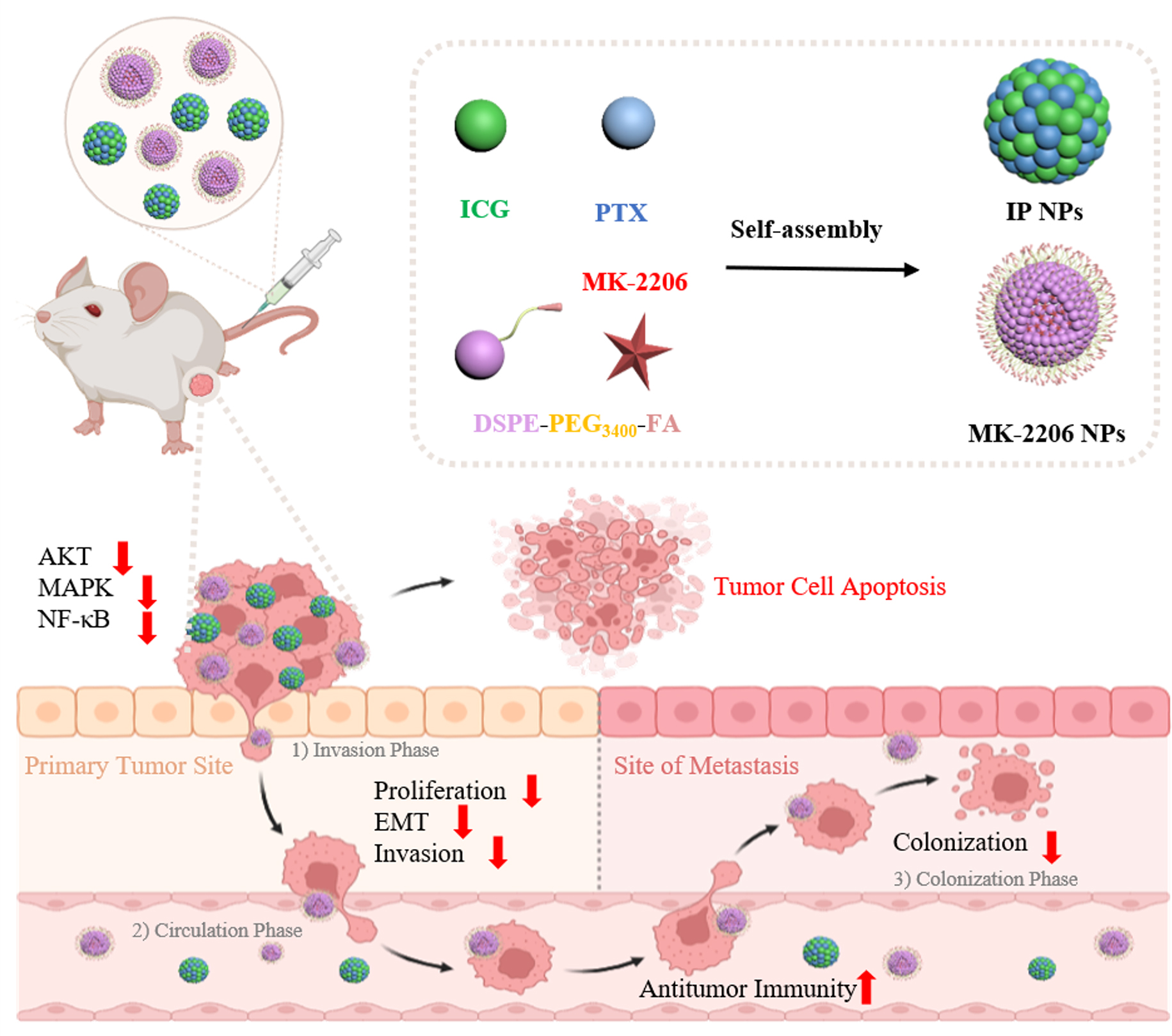

In this study, a plant-derived antitumor drug PTX and commonly used photosensitizer ICG, were applied to prepare carrier-free nanoparticle (IP NPs). During the formation of IP NPs, ICG could be combined with PTX by π-π stacking to increase its solubility. Therefore, to explore the optimal formula for IP NPs formation, different mass ratios (1: 2, 1: 1 and 2: 1) of ICG to PTX were first evaluated. As displayed in Fig. 2A and B, the size distribution of IP NPs was measured, when the ratio of ICG to PTX is 1: 1, IP NPs have a smaller average diameter of 144.40 ± 0.78 nm and polydispersity index (PDI) of 0.27, increasing the mass of PTX or ICG would increase the particle size and PDI of the nanoparticles. The average diameter of IP NPs (1:2) was 153.10 ± 1.49 nm (PDI = 0.35), and 194.17 ± 3.87 nm (PDI = 0.38) was obtained in IP NPs with a feed ratio of 2:1. It was worth noting that in Fig. 2C, as the proportion of ICG increased, the zeta potential of IP NPs decreased gradually. The average zeta potentials of IP NPs were 0.296 mV(-30.2 ± 0.46) (1:2), -17.87 mV (1:1), and − 32.47 mV (2:1) respectively. Zeta potential serves as a critical physicochemical parameter for evaluating the colloidal stability and biodistribution profiles of anticancer drug-loaded nanocarriers, directly influencing their systemic circulation kinetics and cellular internalization efficiency [17]. Anionic nanoparticles demonstrate low hemolytic risk and biocompatibility while prolonging blood circulation via reduced protein adsorption and enhanced stability, due to surface charge preventing RES clearance and electrostatic repulsion reducing erythrocyte binding [18]. Therefore, the use of the 1:2 formula ratio is questionable. Transmission electron microscopy (TEM) was applied to further observe the morphology. Obviously, PTX and ICG at the ratios of 1:2 and 1:1 were likely self-assembled into the nanomedicine with uniform morphology and narrow size distribution, which indicated that the sizes of nanoparticles were approximately 100 nm and in uniform spherical shapes with rough surface (Fig. 2D). By using the UV − vis spectrum, the encapsulation rates of PTX were further measured to be 90.48%, 81.83%, and 90.85%, respectively (Figure S1), indicating that the PTX was mostly reserved during the formation of IP NPs with the manner of feeding ratios un-dependent. Thus, IP NPs with the feed ratio of 1: 1 was selected as the optimal formulation for further in vitro and in vivo studies. Then, the stability of IP NPs was further evaluated. As indicated in Figure S2, the average diameters and PDI values of the nanomedicine were found to have no obvious changes within 7 days which were saved in ultrapure water and PBS.
Then, the self-assembly behavior of IP nanoparticles was investigated through UV-vis spectroscopy and DLS analysis under different environmental conditions. As shown in Figure S3B, DLS revealed significant size augmentation in acidic buffer (pH 5.5, simulating endolysosomal compartments) compared to physiological conditions (pH 7.4), indicative of charge shielding through protonation-induced disruption of interpolymeric [19]. Moreover, it was a remarkable pH-response phenomenon for the drug release because it was slightly acidic in the tumor microenvironment and cellular lysosomes [20, 21]. The carrier-free IP NPs maintained structural integrity during systemic circulation, achieving tumor-specific accumulation before undergoing stimuli-responsive disassembly within the tumor microenvironment, thereby eliciting localized oncotherapeutic effects through controlled payload release. Furthermore, with the addition of hydrophobic sodium dodecyl sulfate (SDS), the absorption peaks of IP NPs were observed to be blue-shifted slightly and the width narrowed (Figure S3C). We suspect that the hydrophobic interaction drove the assembly of IP NPs. In addition, sodium chloride (NaCl) had significant influence on the UV − vis spectrum of IP NPs (Figure S3D), which demonstrated the electrostatic interaction played a role during the formation of IP NPs. In consequence, as shown in Figure S3A, the driving forces of IP NPs formation include π-π stacking, hydrogen bonding, hydrophobic interaction, and electrostatic interaction between PTX and ICG molecules.
Subsequently, the cellular uptake capability of IP NPs was evaluated by flow cytometry (FCM) and observed by inverted fluorescence microscope. As shown in Fig. 2E and S4, time-dependent enhancement of cellular internalization was observed through ICG fluorescence quantification, demonstrating equivalent accumulation levels between IP nanoformulations and their free drug counterparts, suggesting that the nanoparticles formed by the incorporation of PTX did not affect the uptake of drugs by cells. In addition, the photothermal performance is an important evaluation criterion similarly. Then, the in vitro photothermal conversion efficiency of IP NPs was examined with an 808 nm laser (1 W/cm2). As shown in Fig. 2F, the temperature of IP NPs (20 µg/mL) in aqueous solution could reach 44 °C in 2 min under NIR irradiation, which is high enough to damage 4T1 cells. According to our previous research, this is a low ICG usage concentration, so IP NPs also has a good photothermal conversion ability [10].
Fig. 2
Characterization of carrier-free IP NPs. The sizes of IP NPs in (A), PDI in (B), zeta potentials in (C), and TEM images in (D). Data are presented as means ± SEM. (n = 3 independent samples). Scale bar: 1 μm (top) and 200 nm (bottom). (E) Flow cytometry analysis of 4T1 cells treated with 2 µg/mL IP NPs or free ICG for 2, 4 and 8 h. (F) Comparison of the photothermal conversion behaviors of IP NPs at different concentrations under 808 nm laser irradiation (1 W/cm2). The heating and cooling curve is on the left and the infrared image is on the right
The assembly of IP NPs enhances drug responsivenessAfter the above explorations, we had verified the good extracellular physicochemical properties of IP NPs, then we investigated the in vitro synergistic photo-chemo cytotoxicity of IP NPs against mouse mammary carcinoma cell line 4T1. The results of MTT test were shown in Fig. 3A, the cell viability of the Control group was set to 100%, and the cell viability of ICG was still stayed at a higher level (above 80%), which could be considered as non-toxic to cells in this concentration range (0–10 µg/mL). However, the curves of the laser irradiation groups and their respective non-laser irradiation groups almost coincided, revealing the phototherapy of ICG stimulated by NIR hardly had any effect due to the low concentration. Referring to Fig. 2F, at the ICG concentration of 10 µg/mL, photothermal heating could hardly reach 40 ℃ within 5 min, so the concentration of ICG was too low to produce sufficient photothermal effect to cause tumor cell damage. It had been recognized that the assembly formation of IP NPs significantly enhanced the cytotoxicity of PTX at such low drug concentrations, resulting in that free PTX exhibited medium cell viability (60%) but dramatically decreased cell viability (20%) in IP NPs groups. This in vitro study confirmed IP NPs’ concentration-dependent cytotoxicity and their ability to enhance PTX efficacy in tumor cells. Given the greater complexity of the in vivo environment, where the chemotherapeutic effects of IP NPs are limited, subsequent animal experiments are needed to evaluate their combined chemo-phototherapy effects.
To further exclude the synergistic effect of ICG and PTX, we contrasted IP NPs with a mixture of ICG and PTX. To our astonishment, the addition of free ICG hardly changed the cytotoxicity of pure PTX, while the cytotoxicity of IP NPs group was significantly higher than that of the “free ICG + PTX” group (Fig. 3B). This implies that it is the assembly of nanoparticles that enhances the cytotoxicity rather than the synergistic effect of ICG and PTX. PTX and ICG mediated chemotherapy and photothermal therapy respectively to promote tumor cell apoptosis. Therefore, we examined the cell apoptosis rate induced by 5 µg/mL free ICG, free PTX and IP NPs by flow cytometry. As indicated in Fig. 3C and D, the “control” and “ICG” groups only induced a low proportion of apoptosis (12.01% and 9.23%) in 4T1 cells after 24 h of incubation. Higher percentage of apoptotic cells (50.4%) of “PTX” group was observed, which attributed to the inhibitory effect of paclitaxel on cell mitosis and thus on facilitating cell apoptosis. In addition, apoptotic cells induced by IP NPs was significant increase, that caused a majority of apoptotic cells (74.10%), suggesting a great potential of this self-assembly strategy for tumor therapy without the help of photodynamic and photothermal forces.
In addition, the expressions of related representative genes in 4T1 cells could reflect the common tumor cell signaling pathways affected by IP NPs were further examined by RT-qPCR (Figure S5). However, the results revealed that, after treatment with 5 µg/mL of IP NPs, the expression levels of PI3K, AKT3, ERK, and JNK genes were significantly increased, and showed a certain degree of concentration dependence. In tumor cells, the function of JNK branch of MAPK pathway may be abnormally regulated, which may activate anti-apoptotic proteins or inhibit pro-apoptotic proteins, thereby helping tumor cells to escape apoptosis. The ERK branch of the MAPK signaling pathway plays a key role in the process of tumor cell proliferation and the activated ERK can regulate the reorganization of cytoskeletal protein and the expression and activity of matrix metalloproteinases (MMPs) to promote tumor cell migration and invasion. The AKT pathway is an important regulatory pathway for tumor cell survival, and activation of this pathway promotes abnormal tumor cell proliferation and differentiation, and the overexpression of AKT3 may be associated with the aggressiveness and poor prognosis of breast cancer. In addition, the activation of the AKT pathway can make tumor cells resistant to chemotherapy drugs and maintain tumor cell survival by inhibiting apoptosis. Thus, we verified the mRNA expression of P-gp gene in 4T1 cells, it was significantly up-regulated by the IP NPs, which was relieved only when the concentration reached 10 µg/mL. As a typical substrate of PTX, transmembrane transport protein P-gp (P-glycoprotein) can bind to entered PTX molecules and then transport them outside the cell through its transmembrane domain [22].
IP NPs could not effectively inhibit tumor related pathways in 4T1 cells and cause the resistance of tumor cells to chemotherapeutic drugs, implying that the therapeutic strategy of combining molecular targeted drugs might reduce the viability and drug resistance of tumor cells, enhance the comprehensive efficacy of anti-tumor drugs, thus could improve the biological safety by reducing the dose of IP NPs used.
Fig. 3
Cytotoxicity and proapoptotic ability of IP NPs. Cell viability of 4T1 cells after incubation with different concentrations (0–10 µg/mL) of free ICG, free PTX, or IP NPs in dark or under laser irradiation in (A), and comparison of the IP NPs and the mixture of ICG and PTX in the same concentration in (B). L stands for near-infrared laser irradiation (808 nm, 1 W/cm2). (C) Flow cytometry detection apoptosis of 4T1 cells what were incubated with 2 µg/mL of free ICG, free PTX or IP NPs for 24 h, and the results were quantitative analyses in (D)
MK-2206 NPs has the potential to inhibit the proliferation and metastasis of tumor cellsAs a critical proto-oncogenic kinase in the PI3K/AKT/mTOR axis, AKT orchestrates pleiotropic cellular processes including apoptotic suppression, proliferative activation, and metabolic reprogramming through downstream effector modulation [23]. Emerging clinical evidence demonstrates enhanced progression-free survival rates through synergistic integration of AKT inhibitors with front-line chemotherapeutic regimens in breast cancer cohorts harboring pathway-specific genomic alterations [13, 24]. Clinical trials have been reported the AKT inhibitor MK-2206 and standard taxane- and anthracycline-based neoadjuvant therapy improved response in patients with breast cancer in the I-SPY 2 trial [25]. Therefore, targeting AKT and its regulators may provide a feasible solution for the resistance of chemotherapy and cell metastasis.
To increase internalization of tumor cells, the liposome nanoparticles MK-2206 NPs were fabricated through nanoprecipitation approach. The surface of the MK-2206 NPs was functionalized by FA, which helps to deliver the MK-2206 on the site of the breast cancer cells only in a target-specific manner because of the overexpression of FA receptors on the cell surface (Fig. 4A). MK-2206 NPs incorporated with Cy7 were able to emit fluorescence and could be used as fluorescence probes for cell photoimaging to judge the internalization of nanoparticles. As shown in Fig. 4B and C and S6, efficient cellular uptake of MK-2206 NPs in 4T1 cells and 3T3 cells was investigated through inverted fluorescence microscope. After 2 h of incubation with MK-2206 NPs with Cy7, scarcely any fluorescent signals of Cy7 were merged with 3T3 cells, however, clearly fluorescent signal of Cy7 could be seen entering into the cells. As compared to non-neoplastic cells, MK-2206 could recognize tumor cells faster and be engulfed by them. After a longer incubation with the nanoparticles (4 and 8 h), obviously had stronger Cy7 signal in two kinds of cells, nonetheless, 4T1 cells still showed more nanoparticle uptake, indicating that the cellular uptake of nanoparticles was in time-dependent manner and FA modification could enable effective targeted delivery of tumor cells. Then, the cytotoxicity and ability to inhibit AKT activation after cellular uptake of nanoparticles need to be further explored. It has been explored in previous experiments that, within a certain concentration range, MK-2206 treatment of 4T1 cells for 24 h did not have much effect on inhibition of AKT activation in cells (Figure S7). Therefore, the 48 h-incubation with MK-2206 NPs was used in subsequent experiments. As shown in Fig. 4D, the effect of MK-2206 NPs on the cell viability of 4T1 cells was also concentration-dependent, the cell viability was more than 80% when the concentration of nanoparticles was 1 µM and below, that is, the toxicity of MK-2206 NPs to 4T1 cells was negligible within this concentration range. In addition, at the concentration was higher than 10 µM, cell viability was reduced to less than 50%, therefore, the three concentrations (1, 5 and 10 µM) were selected to continue to explore the inhibitory effect of AKT, and the expression of AKT-related proteins was analyzed by western blotting analysis (Fig. 4E-G and S8). As expected, MK-2206 NPs (5 and 10 µM) could inhibit the phosphorylation of AKT with little effect on protein expression. Meanwhile, the EdU staining proliferation kit was used to investigate the effects of MK-2206 NPs on 4T1 cells proliferation to further intuitively show the effect of the inhibition of AKT. Proliferative nuclei were specifically labeled through EdU incorporation, with S-phase cells exhibiting distinct Alexa Fluor 555-tagged fluorescence signals detectable via multispectral imaging. As indicated in Fig. 4H-I, at 24 h of incubation, none of the three concentrations of MK-2206 NPs affected cell proliferation, until 48 h after incubation, MK-2206 NPs (5 and 10 µM) obviously reduced the proportion of proliferating cells. Thus, MK-2206 NPs could effectively inhibit the proliferation of tumor cells, which was consistent with the previous experimental results.
During metastatic progression, neoplastic cells undergo sequential pathophysiological transformations: initial dissociation from primary tumor matrices followed by stromal invasion and subsequent intravasation into adjacent vasculature. This migratory transition is molecularly orchestrated through EMT, a dynamic pathobiological process characterized by transcriptional repression of epithelial markers (e.g., E-cadherin) and concomitant upregulation of mesenchymal phenotypes (e.g., N-cadherin, vimentin) [26]. EMT activation confers malignant cells with enhanced tissue-invasive competence through cytoskeletal reorganization and protease-mediated extracellular matrix degradation. Therefore, E-cadherin and Vimentin can be used as two markers of EMT to show the metastatic ability of tumor cells [27]. We following detected the expression of E-cadherin and Vimentin by RT-qPCR assay, and the results are shown in Fig. 4J. At 24 h of incubation, MK-2206 NPs almost did not affect the expression of the two proteins in cells within the certain concentration range, while at 48 h of incubation, 5 µM and 10 µM MK-2206 NPs significantly upregulated E-cadherin expression and downregulated Vimentin expression, that is in contrast to the EMT process. These data confirmed the MK-2206 NPs have the potential of faster uptake by tumor cells with folate receptor overexpression on the cell membrane surface, effectively inhibits AKT activation, inhibits cell proliferation, and have the potential of inhibiting EMT to decreases cell invasiveness.
Fig. 4
The mechanism of MK-2206 NPs’ ability to inhibit tumor cell metastasis. (A) Schematic illustration of MK-2206 NPs targeting FA receptor overexpressed breast cancer cells. (B) Fluorescence images of cellular uptake by 4T1 cells or 3T3 cells for 2 h, and the quantitative analysis of mean fluorescence intensity was shown in (C). (D) Cell viability of 4T1 cells after incubation with different concentrations (0–10 µM) of MK-2206 NPs. Quantitative analysis of relative protein expression ((E) pAkt/β-actin, (F) pAkt/Akt, and (G) Akt/β-actin) in 4T1 cells incubated with different concentrations of MK-2206 NPs for 48 h. Data are presented as means ± SEM. (n = 3). (H) Fluorescence images of the EdU assay show proliferating cells, and the quantitative analysis in (I). Scale bar: 100 μm. (J) RT-qPCR experimental results. E-cadherin and Vimentin gene expression levels in cells, n = 4. The statistical significance was analyzed by one-way ANOVA with a Tukey post hoc test. p-value: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
IP NPs and MK-2206 NPs synergistically inhibited the metastasis of tumor cells in vitroFor better validation of the potency of combination therapy of IP NPs and MK-2206 NPs, cell wound scratch assay, trans-well cell migration assay, and vein epithelial cell adhesion assay were further applied to visualize the ability of the tumor to metastasize. Firstly, MTT assay showed that the two kinds of nanoparticles had synergistic effects on cell damage (Figure S9). Refer to Figs. 3A and 4F, we chose separately the concentration of IP NPs (1 µg/mL) that produced cell viability at 50% and the concentration of MK-2206 NPs (5 µM) that produced an effect on pAKT expression. Cell wound scratch assay was applied to verify preliminarily the effect of synergistic use of IP NPs and MK-2206 NPs on cell migration. As shown in Fig. 5A-C, a small green dot represented a living 4T1 cell in images, compared to the control group, negligible inhibition of cell migration was found in “ICG” group. Obviously, IP NPs showed higher potential against 4T1 cells migration than PTX, and this effect become more pronounced with the synergy of MK-2206 NPs in “IP/M” group. To our surprise, the cells became more sparsely arranged after incubation with MK-2206 NPs and IP NPs, and could be observed that there were many voids in the cell layer, a part of the cells died and no longer adhered to the bottom of petri dish, indicating that the combined therapeutic approach of MK-2206 pathway suppression and carrier-free nanoparticle assembly significantly enhances cytotoxic effects in 4T1 cells, effectively inhibiting both proliferation and migration through synergistic mechanisms.
The migration abilities of 4T1 cells incubated with different drugs were further evaluated by Trans-well cell migration assay. For the analysis, the number of 4T1 cells that penetrated the Matrigel-coated polycarbonate membrane in the “PTX” and “MK-2206 NPs” groups were 51.00% and of 56.75% that in the control group benefit from the damage effect of the drug on tumor cells. “IP NPs” group exhibited strong invasion-inhibition abilities (15.00%), particularly, the combined effect of IP NPs and MK-2206 NPs almost completely inhibited the metastasis of tumor cells (Fig. 5D-F). During metastatic progression, tumor cells detach from primary sites, intravasate into the circulatory system, and persist as circulating tumor cells (CTCs), representing a critical phase in cancer dissemination. The adhesion of CTCs to ECs in a premetastatic niche is essential for metastasis formation [28]. Thus, we investigated whether IP NPs and MK-2206 could hinder the adhesion of tumor cells to ECs in the pre-metastatic site. HUVECs were pre-cultivate to mimic the ECs in the niche, and the number of 4T1 cells that adhered to IP/M-treated HUVEC monolayer was significantly reduced compared with those in other groups (Fig. 5G-H). Then, we studied the expression of main cell adhesion molecule ICAM-1 on HUVECs and 4T1-GFP cells in the cell plates by qRT-PCR to explore the mechanism. ICAM1 facilitates metastatic progression through modulation of cell cycle regulation and stemness maintenance pathways, representing a promising molecular target for inhibiting early-stage metastasis [29]. As shown in Fig. 5I, the significantly reduced expression of ICAM1 revealed that IP NPs and MK-2206 NPs were able to effectively inhibit adhesion between ECs and tumor cells, that is consistent with the results of the in vitro simulated adhesion assay described above.
Fig. 5
Combined anti-metastasis effect of IP NPs and MK-2206 NPs. Schematic illustrations of wound-healing assay (A), trans-well cell migration assay (D), and epithelial cell adhesion assay (G). (B), (C) Cell wound healing rate of 4T1 cells (labeled calcein-AM) incubated with nanoparticles (the concentration of IP NPs was 1 µg/mL and the MK-2206 NPs was 5µM, and the same below), n = 3, scale bar: 250 μm. (F) Relative quantitative cell migration of 4T1 cells, n = 4 and (F) representative images. Scale bar: 100 μm. (H) Images of 4T1-GFP cells adhered to HUVECs. Top photo, bright field; middle photo, fluorescent field. Scale bar: 200 μm. The white arrow represents the void created in the dense layer of HUVECs cells at the bottom. (I) RT-qPCR experimental result of ICAM-1 gene expression levels in the co-culture mixture of 4T1 cells and HUVECs cells, n = 4. The statistical significance was analyzed by one-way ANOVA with a Tukey post hoc test. p-value: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
IP NPs and MK-2206 NPs synergistically regulate cellular pathwaysWe have confirmed that IP NPs and MK-2206 NPs could efficiently hinder tumor cell metastasis by reducing cell invasiveness and adhesion from three cell experiments. Furthermore, in order to explore the underlying mechanism, we studied the expression of some main cell molecules of tumor-related pathways. In Sect. 3.2, we have briefly presented that activation of MAPK and AKT pathways contributes to tumor cell survival. The MAPK and AKT pathways are closely related and often cooperate with each other in tumor cells. They have a cross-activation mechanism and jointly promote the occurrence and development of tumors. In certain cases, the MAPK pathway can also activate the AKT pathway. For instance, the activation of extracellular signal-regulated kinase (ERK) can enhance the activity of PI3K by phosphorylating the regulatory subunit of PI3K, which in turn indirectly activates the AKT pathway. In addition, the NF-κB pathway also plays an important role in the process of tumorigenesis, development and metastasis. Phosphorylated AKT can directly or indirectly activate the NF-κB pathway in a variety of ways. When AKT phosphorylates IκB Kinase (IKK), IKK is activated, which in turn phosphorylates IκB protein. The phosphorylated IκB protein is recognized and degraded by the ubiquitin-proteasome system, resulting in the release of NF-κB from the cytoplasm and translocation to the nucleus, initiating the transcription of related genes. Mutation or abnormal activation of the AKT pathway can lead to the activation of the NF-κB pathway, which in turn promotes the proliferation, survival and migration of tumor cells. Schematic diagram of the relationships between various pathways and their effects on cell survival, growth, proliferation, apoptosis, and EMT was shown in Fig. 6A. The combined treatment of IP NPs and MK-2206 NPs was conducted to investigate the impact of the drugs on cell signaling pathways.
Therefore, the expression of AKT on 4T1 cells was analyzed by Western blot. As depicted in Fig. 6B, following treatment with concentrations of nanoparticles (1 µg/mL IP NPs, 5 µM MK-2206 NPs) for 48 h, the expression of pAKT in the ICG NPs group, the MK-2206 NPs group, and the IP/M group were significantly decreased, indicating the activation of AKT proteins were prominently inhibited. It might be that the concentrations of both nanoparticles are too high to block the synergistic effect. Thus, the lower concentration of nanoparticles (0.5 µg/mL IP NPs, 1 µM MK-2206 NPs) was utilized for subsequent treatments. As shown in Fig. 6C, the inhibitory effect of ICG NPs on AKT protein activation was nearly negligible, and the effect of MK-2206 NPs was also not very obvious, which was in accordance with the previous exploratory experiments, thus, 1 µM MK-2206 NPs was unable to inhibit the activation of AKT protein, that was consistent with the previous results. However, when the two nanomedicines at low concentrations were used for combined treatment, the inhibition of AKT protein activation was relatively pronounced, revealing the favorable outcome of the synergistic treatment in regulating the AKT pathway and reducing the dosage of anticancer drugs used.
Similarly, lower concentrations (0.5 µg/mL IP NPs, 1 µM MK-2206 NPs) of nanoparticles have the potential to synergistically inhibit the activation of ERK, IκB-α, and NF-κB proteins in tumor cells and the synergistic effect of the two nanoparticles was not obvious at higher concentrations (1 µg/mL IP NPs, 5 µM MK-2206 NPs) (Fig. 6B-C). It is remarkable that, pure PTX significantly increased the protein expression of pNF-κB especially at lower dose, that is, significantly promoted the activation of NF-κB, while the other groups did not have this phenomenon. And the synergistic inhibition effect of IP NPs and MK-2206 NPs was more obvious, indicating that the use of pure drugs may stimulate the enhancement of drug resistance of 4T1 cells to ensure their survival, while IP NPs and MK-2206 NPs may reduce the drug resistance of cells by inhibiting the activation of pathways, thereby improving the toxicity of drugs to tumor cells. In addition, IP NPs, MK-2206 NPs, and IP/M three groups all significantly promoted the degradation of IκB-α at higher dose. Theoretically, phosphorylation of IκB-α would lead to ubiquitination and degradation of IκB-α. It facilitates the translocation of NF-κB protein into the nucleus. This was in contrast to the decrease of pNF-κB and pIκB-α protein expression in the experimental results. However, it is worth noting that in Fig. 6B, the expressions of these four proteins and their respective phosphorylated proteins in the NF-κB pathway were significantly decreased, indicating that the NF-κB pathway was inhibited as a whole.
The results presented above indicated that the synergy between IP NPs and MK − 2206 NPs effectively regulated the AKT, MAPK, and NF - κB pathways in 4T1 cells, suggesting the potential to inhibit the tumor cell survival and metastasis. Once these pathways were modulated, the downstream effector factors were the ones that actually exerted the effects. Therefore, we measured the mRNA expression levels of Vimentin, E-cadherin, Bax and Bcl-2 by RT-qPCR assay. The results are depicted in Fig. 6D. Both the IP NPs group and the IP/M group exhibited a decrease in Vimentin expression. Regarding E-cadherin expression, drug treatment significantly enhanced it, and the effect of IP NPs was notably stronger than that of free PTX, with a pronounced synergistic effect. Evidently, the combined treatment of IP NPs and MK-2206 NPs could more effectively inhibit tumor cell metastasis by suppressing EMT (Epithelial - Mesenchymal Transition). Moreover, the viability and state of tumor cells are also crucial factors influencing their metastatic capacity. From the results we found that, the first five groups had no significant impact on the expression of the pro-apoptotic factor Bax in cells. However, the combined treatment group (IP/M) significantly upregulated the mRNA level of Bax. For the anti-apoptotic factor Bcl-2, which counteracts Bax, the last four groups all showed a significant downregulation, and the IP/M group had an even lower expression level. In conclusion, the combined treatment of IP NPs and MK-2206 NPs can better inhibit tumor cell metastasis by reducing the invasiveness and promoting apoptosis of tumor cells.
Fig. 6
Regulation of cellular pathways (Akt, MAPK, and NF-κB) by IP NPs and MK-2206 NPs. (A) Schematic diagram of the relationships between various cell pathways and their effects on cell survival, growth, proliferation, apoptosis, and EMT. Western blotting analysis of the 4T1 cells treated by higher dose nanoparticles (1 µg/mL IP NPs and 5 µM MK-2206 NPs) (B) and lower dose nanoparticles (0.5 µg/mL IP NPs and 1 µM MK-2206 NPs) (C) for 48 h. Relative expression quantification of various proteins on the right, n = 3. (D) RT-qPCR experimental results. EMT-related (Vimentin and E-cadherin) and apoptosis-related (Bax and Bcl-2) gene expression levels in cells treated by lower dose nanoparticles (0.5 µg/mL IP NPs and 1 µM MK-2206 NPs) for 48 h, n = 4. The statistical significance was analyzed by Student’s t-test. p-value: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
IP NPs and MK-2206 NPs synergistically inhibited the tumor growth and metastasis in liver and lungOwing to the satisfactory ability to regulate cellular pathways and inhibition of tumor cell metastasis, to determine whether the Carrier-free nano-integrated strategy for synergetic AKT targeted therapy can serve as a potential therapy to chemotherapy and phototherapy, the in vivo antitumor efficacy was next evaluated on 4T1 subcutaneous tumor model. Firstly, the biodistribution of IP NPs was detected by fluorescence imaging. As displayed in Fig. 7A-C, systemic administration of IP NPs resulted in comprehensive biodistribution, facilitated by circulatory transport to multiple organ systems. And IP NPs had significantly longer retention time in vivo, while ICG was metabolized faster. The IP NPs demonstrated significantly extended plasma residence time relative to free ICG, potentially due to enhanced biocompatibility achieved through molecular self-assembly. Notably, 12-hour post-injection analysis revealed significant fluorescence accumulation in tumor tissues from IP NP-treated subjects, demonstrating enhanced tumor retention through nanoformulation.
Fig. 7
Biological distribution of IP NPs. (A) Schematic diagram of tail vein injection (5 mg/kg free ICG or IP NPs), in vivo imaging and in vitro organ imaging. (B) Time-lapse fluorescent images of 4T1 breast cancer-bearing mice and quantitative analysis in (C), and the yellow circle indicates the location of the tumor, n = 3. (D) Fluorescent images of tumors and major organs isolated from mice receiving treatments for 12 h and quantitative analysis in (E), n = 3
Firstly, a certain amount of 4T1-luc cells was injected into the tail of non-tumor-bearing mice in each treatment group to allow systemic dissemination. The fluorescence signals from luciferin indicated that the combination of IP and MK-2206 nanoparticles partially inhibited tumor cell metastasis and lung colonization. However, due to the absence of ICG-mediated phototherapy, the metastasis inhibition effect was limited (Figure S11). Then, therapeutic efficacy evaluation was conducted in orthotopic 4T1 breast cancer models following systemic administration of the experimental agents. As shown in Fig. 8A and S12A, the tumor volumes were monitored every 3 days after various treatments. The tumor growth curves of different groups were drawn as displayed in Fig. 8B-E and S12B-D. In these examples, the tumor volumes of the Free PTX, MK-2206 NPs, IP/M, Free ICG + L and IP NPs groups had the comparable growing tendency to those of Control group, illustrating that nanoparticles did not prevent the tumor proliferation at all in the absence of light, and the injection of free ICG could not achieve sufficient local heating to damage tumor cells even if laser irradiation is performed because of insufficient accumulation in the tumor sites (Figure S12E). And IP NPs and IP/M groups could partially inhibit the tumor growth, resulting phototherapy was necessary for in vivo experiments. Thus, IP NPs + L and IP/M + L groups could effectively inhibit the tumor growth with our expectations, and both groups also significantly inhibited tumor cell proliferation that the labeling rate of Ki67 was significantly reduced as shown in Fig. 8F.
Fig. 8
The red circle indicates that the tumor has been completely ablated
However, on the basis of tumor inhibition alone, the synergistic effect of MK-2206 NPs was not obvious. Therefore, we further explore from the aspect of anti-tumor metastasis. On the day 18 of treatment, we collect a subset of tumors and the expression of proteins was determined by Western blot assay. IP/M + L group significantly reduced the ratio of phosphorylated to non-phosphorylated protein amount, that is, inhibited the activation of ERK, JNK, NF-κB and AKT proteins (Fig. 9A and S13). The synergistic effect of IP NPs and MK-2206 NPs with photothermal significantly regulated cell pathways related to proliferation, metastasis, drug resistance and other cellular pathways, demonstrating the potential of IP NPs and MK-2206 NPs in inhibiting tumor metastasis. Then, we counted the number of liver metastases nodules in liver sections and observed the H&E staining sections of lung and liver tissue (Fig. 9B-F and S14). The mean metastatic nodules in the livers of mice treated with control, IP NPs, IP NPs + L, IP/M + L were 26, 16, 1, and 0, respectively. Moreover, obvious inflammatory infiltration was observed in control, IP NPs, and IP NPs + L groups, which reveal the high levels of hepatic inflammation. While the pathogenesis of inflammation involves multiple factors, perivascular inflammatory foci in the hepatic tissues may result from pathophysiological processes triggered by the hematogenous spread of neoplastic cells through endothelial adhesion and cytokine-mediated activation. As time went on, there was preferred to significant parenchymal lung tissue in control and IP NPs groups and both groups had significantly higher liver and lung weights. All these results suggested that the synergistic treatment strategies effectively inhibited the growth, proliferation, and metastasis of tumor cells.
Fig. 9
The anti-metastasis efficacy of IP NPs and MK-2206 NPs in 4T1 orthotropic primary tumor-bearing mice. (A) Relative expression quantification of western blotting analysis of the resected tumor after 18 d of treatment. (B) Typical liver tissues with visualized metastatic nodules (yellow dotted lines) from each group, and representative fields of liver tissues with metastasis areas (yellow arrows and dotted lines) from each group with H&E after 18 and 30 d of treatment. Scale bar: 200 μm. (C) H&E of lung tissues. Scale bar: 100 μm. (D) The number of metastatic nodules in liver. The weight of livers (E) and lungs (F). The statistical significance was analyzed by one-way ANOVA with a Tukey post hoc test. p-value: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001
IP NPs and MK-2206 NPs synergistically inhibited the recurrence of the tumorPrevious studies indicate that tumor tissue damage can generate in situ tumor vaccines, potentially activating anti-tumor immune responses and preventing cancer recurrence. In order to study the effect of synergistic therapy on tumor rechallenge and recurrence, we built a 4T1 tumor rechallenge model, as shown in Fig. 10A. Following identical nanotherapeutic administration as in subcutaneous models, surgical tumor resection was performed, with subsequent tumor cell rechallenge near the surgical site to evaluate therapeutic memory effects. We first demonstrated again the inhibitory effect on subcutaneous tumors in Fig. 10B-D, surprisingly, compared to the control group, all tumors in IP/M + L group were ablated, but a small amount of tumor remained in IP NPs + L group and were surgical resection. The growth curves of the de novo tumors were also shown in Fig. 10D-F, after IP/M + L treatment, the tumors grew significantly more slowly, and the control group had obvious recurrence, this was reflected in the fact that the volume growth rate of the rechallenge tumors was almost the same as that of the inoculated tumors on healthy mice. We also discovered that, in comparison with the control group, the spleens of the latter two groups were considerably smaller as shown in Fig. 10G. This finding suggests that the combination therapy not only inhibits tumor growth but also alleviates the compensatory enlargement of the spleen during this process. Moreover, this serves as both an indicator of biosafety and a signal of a more robust immune capacity of the body. Then, we collected the spleens of mice on the 14th and 28th days after the first treatment to prepare single-cell suspensions, and analyzed the proportion of cytotoxic T lymphocytes (CD8 +) and helper T lymphocytes (CD4 +) within CD3 + T cells in the primary spleen cells by flow cytometry (Fig. 10H and S15-16). The results showed that the content of CD3 + CD4 + T cells and CD3 + CD8 + was the highest after IP/M + L treatment, indicating that the synergistic therapy has the potential of eliciting a strong anti-tumor immune response, this holds critical importance in suppressing tumor metastasis and recurrence.
Effectiveness and safety are both essential factors that determine the clinical application of cancer therapy. During the treatment period,
Comments (0)