
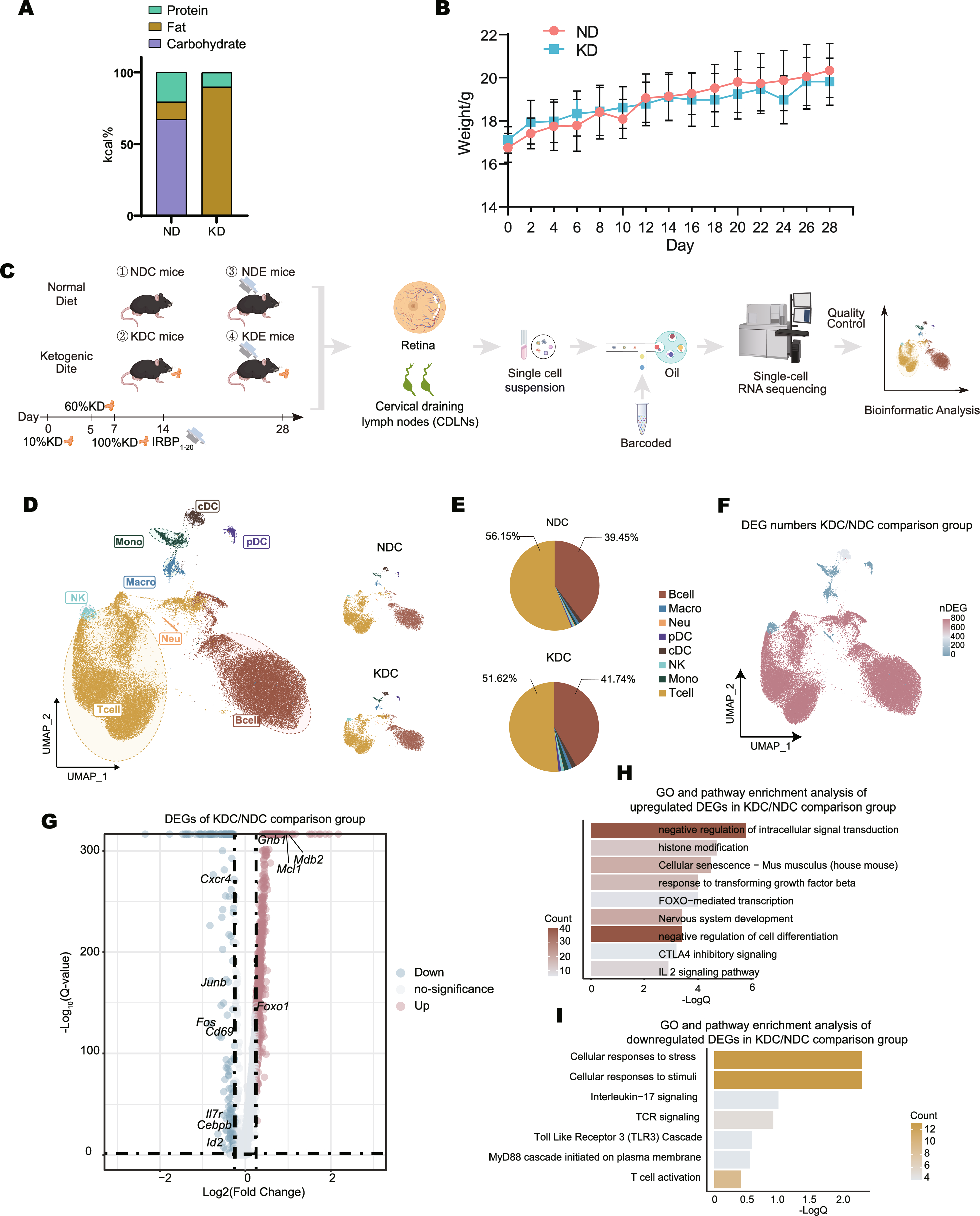
Remember me
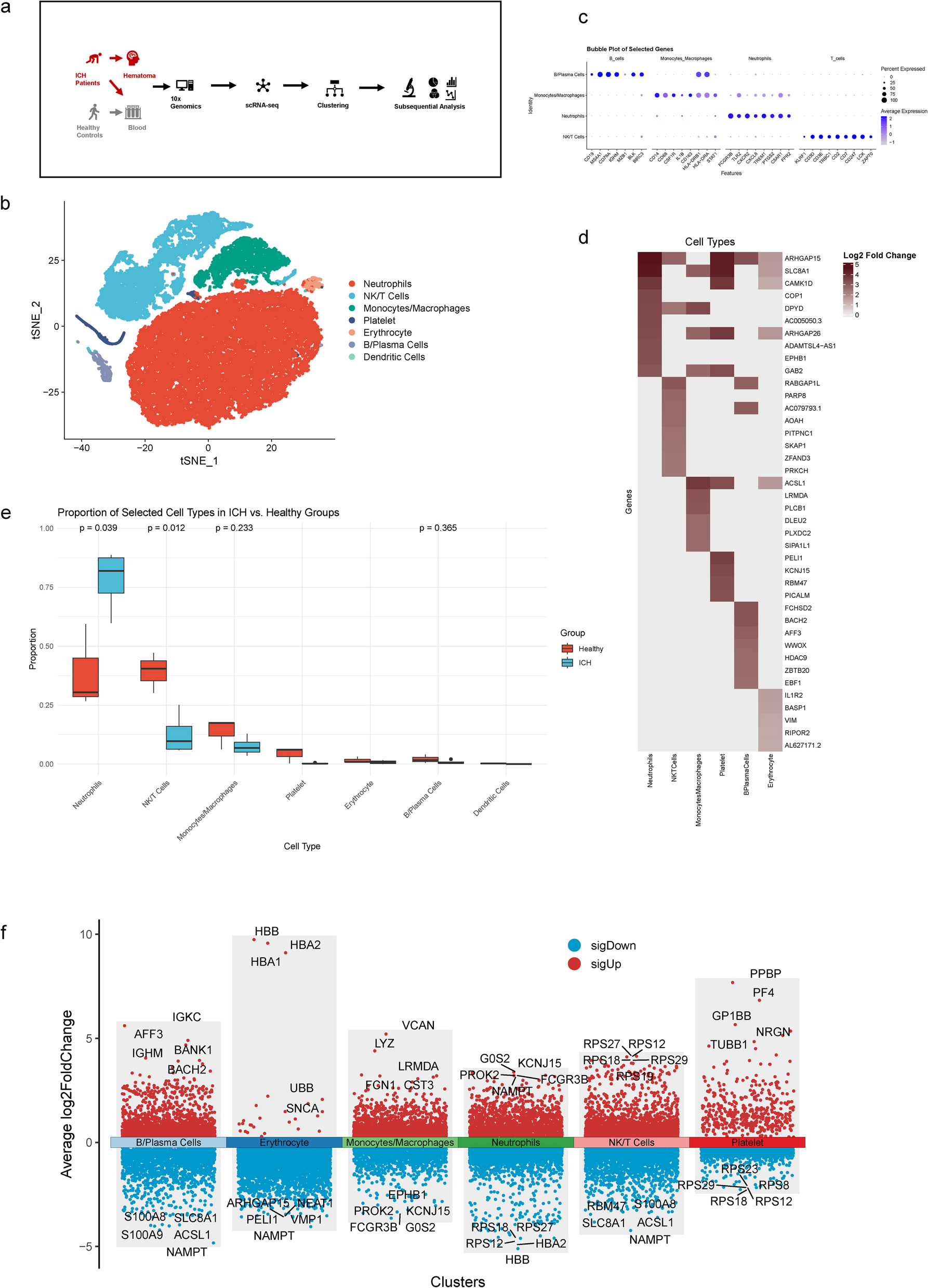


This study enrolled four patients with primary ICH to collect both hematoma and peripheral venous blood samples, with blood samples from three healthy volunteers as control. scRNA-seq was performed on these samples, focusing on immune cells to characterize the immunological profile. Additionally, scRNA-seq data of whole peripheral venous blood cells from three healthy volunteers were obtained to serve as the healthy control group. Baseline information were summarized in (Table S1). We first compared the cellular differences in peripheral venous blood between patients with ICH and healthy controls. Subsequently, we compared the cellular differences between peripheral venous blood and hematoma samples to delineate their immunological characteristics (Fig. 1a).
Fig. 1
Single-cell transcriptomic profiling of venous blood samples derived from ICH patients and healthy controls. a Schematic representation of the study design. b t-Distributed Stochastic Neighbor Embedding (t-SNE) plot of single-cell transcriptomes combining venous blood samples derived from ICH patients and healthy controls. c Bubble plot showing the expression levels of the marker genes in the 4 main immune cell subtypes. d Heatmap illustrating the top up-regulated genes among cell types in ICH patients compared with healthy controls. e Box plots comparing the proportion of selected cell types in ICH patients and healthy controls. f Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each cell subtypes, compared with other cell types in ICH patients
Single-cell transcriptomic profiling of blood samplesWe combined scRNA-seq data from peripheral venous blood samples of ICH patients and healthy controls and performed t-distributed stochastic neighbor embedding (t-SNE) to visualize the transcriptomes of seven major cell types, including four main immune cell types: Neutrophils, NK/T cells, Monocytes/Macrophages, and B/Plasma Cells (Fig. 1b). Dendritic cells were identified exclusively in healthy controls (Figure S1a). The expression levels of marker genes for the four main immune cell types were assessed and visualized, including FCGR3B, TLR2, CXCR2, CXCL8, TREM1, PTGS2, C5AR1, and FPR2 for Neutrophils; KLRF1, CD3D, CD3E, TRBC1, CD2, CD7, CD247, LCK, and ZAP70 for NK/T cells; CD14, CD68, CSF1R, IL1B, CD163, HLA − DRB1, HLA − DRA, and STAT1 for Monocytes/Macrophages; CD19, MS4A1, CD79A, IGHM, MZB1, BLK, and BIRC3 for B/Plasma Cells (Fig. 1c). The distribution of these marker genes was also verified in the t-SNE maps (Figure S1d). The proportions of the seven major cell types between the ICH and healthy control groups were calculated and compared (Fig. 1e). Compared with healthy controls, Neutrophils exhibited elevated proportions (p = 0.039), while NK/T cells showed decreased proportions (p = 0.012) in ICH patients. Monocytes/Macrophages and B/Plasma Cells also exhibited slightly increased proportions. As key mediators, Neutrophils have been reported to play a crucial role in immune and inflammatory processes, as well as in crosstalk with other immune cells, during the early stages of ICH [13]. Additionally, the top five upregulated and downregulated genes for each cell type in ICH patients are displayed (Fig. 1f). Activation of FcγRIIIb (FCGR3B) can lead to the release of inflammatory mediators from neutrophils, thereby participating in the regulation of local inflammatory responses [14]. Cystatin C (CST3) has been infrequently studied in cerebrovascular diseases in relation to monocytes. In cardiovascular diseases, CST3 plays a critical role in enhancing monocyte adhesion to endothelial cells [15]. Differentially Expressed Genes (DEGs) between ICH patients and healthy controls were used to explore major biological functions through Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis, and Gene Set Enrichment Analysis (GSEA) (Fig. 1d; Figure S1e-h). Immune-related biological activities, including GTPase activation, were enhanced, while metabolic-related activities were reduced. Notably, several signaling pathways were significantly activated in venous blood following ICH, including the Neurotrophin, Sphingolipid, Chemokine, MAPK, Fc epsilon RI, and T cell receptor signaling pathways. These changes suggest enhanced biological activities, including the initiation of immune responses, stress responses, apoptosis, and tissue repair.
Cellular interaction analysis in blood samplesCell communication network analysis was conducted on venous blood samples from ICH patients. Among the major cell types, Neutrophils exhibited stronger incoming and outgoing interactions compared to other cell types (Fig. 2a). The contribution of each outgoing communication pattern of secreting cells and incoming communication pattern of target cells was also described (Figure S2a). Compared to healthy controls, venous blood from ICH patients exhibited a decreased number of inferred interactions but increased interaction strength (Fig. 2b). The interaction weights, strength, and numbers among all major cell types were also investigated (Fig. 2c). In ICH patients, Neutrophils, NK/T cells, and Platelets sent signals to all other cell types except Erythrocytes, whereas Monocytes/Macrophages and B/Plasma Cells communicated with all cell types, including Erythrocytes (Figure S2b). The expression levels of key transcription factors were compared between venous blood samples from ICH patients and healthy controls (Fig. 2d). Notably, compared with healthy controls, Human Immunodeficiency Virus Type I Enhancer Binding Protein 2 (HIVEP2) and Class II Major Histocompatibility Complex Transactivator (CIITA) were found to be elevated in nearly all cell types in ICH patients. These factors are essential for regulating immune function and the antigen presentation process during immune responses, supporting the normal functioning of the immune system [16, 17]. Metabolic pathway activity analysis revealed that the TCA cycle, Glycolysis, and Oxidative phosphorylation were significantly reduced in ICH patients (Fig. 2e-g).
Fig. 2
Comprehensive analysis of cellular interactions and metabolic pathways in blood samples derived from ICH patients and healthy controls. a Correlation between incoming and outgoing interaction strength among all pathways across different cell subtypes in blood samples derived from ICH patients. b Comparison of total number of inferred interactions and interaction strength on blood samples between ICH patients and healthy controls. c Number of interactions and interaction weights/strength among different cell subtypes in blood samples derived from ICH patients. d Expression levels of transcript factors across various cell types in ICH patients compared with healthy controls. e Comparison on metabolic pathway activity strength between ICH patients and healthy controls. f Comparison on metabolic pathway activity strength among different cell types from ICH patients. g Comparison on metabolic pathway activity strength among different cell types from healthy controls
Transcriptomic profiling of hematoma samples using scRNA sequencingscRNA-seq data from hematoma and peripheral venous blood samples of ICH patients were subsequently collected to compare transcriptomic profiles and characterize immunological features. Using t-SNE, we identified seven major cell types, including four key immune cell subsets: neutrophils, NK/T cells, monocytes/macrophages, and B/plasma cells (Fig. 3a; Figure S3a). From venous blood to hematoma, the proportion of neutrophils and monocytes/macrophages slightly decreased, while the proportion of NK/T cells increased (Fig. 3b). However, none of the differences in the proportions of cell types between blood and hematoma reached statistical significance. The proportion of neutrophils in the hematoma was higher than that of other subtypes, suggesting a predominant role in the immune and inflammatory processes during the early stage of ICH. The top upregulated genes in different cell types within the hematoma are also shown (Fig. 3c). GO enrichment analyses identified key biological functions in hematoma, compared with blood(Figure S3f). The Type II interferon-mediated signaling pathway was significantly activated and is known to be crucial for T cell activation, enhancing immune responses by stimulating other cell types involved in the immune response [18].
Fig. 3
Single-cell transcriptomic profiling of hematoma and venous blood samples derived from ICH patients. a t-SNE plot of single-cell transcriptomes combining hematoma and venous blood samples derived from ICH patients. b Comparison on the proportion of selected cell types in hematoma and blood samples. c Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each cell subtypes, compared with other cell types in hematoma samples
Cellular interaction analysis in hematoma samplesWe further performed cell–cell communication network analysis on hematoma samples. Neutrophils exhibited the highest incoming and outgoing interaction strength among all seven cell types in hematoma samples (Fig. 4a). The contribution of each outgoing communication pattern from secreting cells and incoming communication pattern to target cells was also described (Figure S4a). An increase in the number of inferred interactions and a decrease in interaction strength were observed in hematoma samples compared to venous blood samples (Fig. 4b). In hematoma samples, Erythrocytes sent signals to all other cell types (Fig. 4c; Figure S4b). C-X-C motif chemokine ligand (CXCL) and Adhesion G protein-coupled receptor E5 (ADGRE5) were the major outgoing and incoming signals for neutrophils in hematoma samples, playing critical roles in neutrophil adhesion, migration, inflammatory regulation, interaction with the complement system, transendothelial migration, BBB disruption, and subsequent tissue repair processes [19, 20] (Fig. 4d). The chemokine receptor 2 (CXCR2) axis was enhanced to facilitate the mobilization of neutrophils from the circulation to the injury site [21]. HIVEP2 and CIITA were significantly upregulated transcription factors in nearly all cell types within the hematoma, compared with blood (Fig. 4e). Metabolic pathway activity analysis revealed that, compared to venous blood samples, TCA cycle was reduced in hematoma samples, whereas the activities of glycolysis/gluconeogenesis and oxidative phosphorylation were increased, suggesting enhanced reactive oxygen species production and impaired energy metabolism due to mitochondrial dysfunction and insufficient energy supply (Fig. 4f-g). Besides, Glutamate metabolism activity was enhanced in hematoma samples, compared with blood samples, indicating excitotoxicity due to accumulation of glutamate, particularly in Glia [22].
Fig. 4
Comprehensive analysis of cellular interactions and metabolic pathways in hematoma samples derived from ICH patients. a Correlation between incoming and outgoing interaction strength among all pathways across different cell subtypes in hematoma samples. b Comparison of total number of inferred interactions and interaction strength between hematoma and blood samples. c Number of interactions and interaction weights/strength among different cell subtypes in hematoma samples. d Outgoing and incoming signaling patterns of each pathways across different cell subtypes in hematoma samples. e Expression levels of transcript factors across various cell types in hematoma compared with blood samples. f Comparison on metabolic pathway activity strength between hematoma and blood. g Comparison on metabolic pathway activity strength among different cell types in hematoma
Immune cell subtypes in blood and hematoma samplesTo gain a deeper understanding of the transcriptomic alterations in major immune cell types from venous blood and hematoma samples in ICH patients, reclustering was performed for each immune cell type, including Neutrophils, NK/T Cells, and Monocytes/Macrophages. In blood samples, Neutrophils were divided into four sub-clusters (Neu0, Neu1, Neu2, Neu3), identified using the Uniform Manifold Approximation and Projection (UMAP) method (Fig. 5i-a). For Neu1-3, the top upregulated genes included CNTNAP3, DUSP6, EGR1, ADAMTSL4 − AS1, and TUBA1A for Neu1 (a potential pro-inflammatory phenotype [23, 24]); KYNU, SDK1, FGF13, IL18RAP, and PHACTR1 for Neu2(a potential anti-inflammatory phenotype [25, 26]); and AL139042.1, IL1R1, IL18R1, AC007271.1, and IL1R2 for Neu3((a potential regulatory phenotype [27]) (Fig. 5i-b). Additionally, the proportions of each Neutrophil sub-cluster were compared between ICH patients and healthy controls. In healthy controls, Neu3 predominated, with a significantly higher proportion than Neu0. In ICH patients, the proportions of Neu1-3 were similar, with a much lower proportion of Neu0 (Fig. 5i-c). For transcription factors, HIVEP2 was upregulated in Neu0-3, while most transcription factors were only upregulated in Neu1-3, including AIRE, CIITA, NFYC, ELF2, SPI1, DMTF1, ATF1, and RCOR2 in ICH patients compared to healthy controls (Fig. 5i-d). Among them, Spi-1 Proto-Oncogene(SPI1) was reported to be crucial in differentiation and cell cycle arrest of Neutrophils [28]. Furthermore, the biological functions of Neutrophils in ICH patients were investigated using GO and KEGG analyses (Figure S5a-c). The immune response process, including the regulation and activation of signaling pathways, was activated in ICH patients. The MAPK signaling pathway, chemokine signaling pathway, and NOD-like receptor signaling pathway were significantly upregulated in Neutrophils, suggesting activation of autophagy in addition to immune responses [29].
Fig. 5
Compositional analysis of neutrophils cell types. i.Blood samples. a The uniform manifold approximation and projection (UMAP) projection showed the subtypes and distribution of Neutrophils. b Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each Neutrophil subtypes. c Comparison the proportion of Neutrophil subtypes from blood samples between ICH patients and healthy controls. d Expression levels of transcript factors across Neutrophil subtypes in hematoma, compared with blood samples. e Top 10 up-regulated genes differentially according to pseudotime. ii.Hematoma samples. a The uniform manifold approximation and projection (UMAP) projection showed the subtypes and distribution of Neutrophils. b Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each Neutrophil subtypes. c Comparison the proportion of Neutrophil subtypes blood and hematoma samples. d Expression levels of transcript factors across Neutrophil subtypes in hematoma, compared with blood samples. e Top 10 up-regulated genes differentially according to pseudotime
In hematoma samples, Neutrophils were reclustered into three subtypes (Neu0, Neu1, Neu2) using the UMAP method (Fig. 5ii-a). In Neu1 from hematoma samples, the top five upregulated genes were FOS, HBB, SSH2, IL1R2, and SORL1, whereas in Neu2, the top five upregulated genes were AC119674, PLPP3, PLAU, PI3, and VEGFA (Fig. 5ii-b). Neu1 and Neu2 constituted the predominant neutrophil sub-clusters in both hematoma and blood samples from ICH patients. Neu1 was slightly more prevalent in hematoma samples, while Neu2 was more abundant in blood samples (Fig. 5ii-c). Among the transcription factors, HIVEP2 was upregulated in Neu0-2, while CIITA was upregulated in both Neu1 and Neu2. Additionally, ID4 was upregulated in Neu1, and GTF3A was upregulated in Neu2 (Fig. 5ii-d).
Given the higher proportion of neutrophils compared to other cell types in both hematoma and blood samples, pseudotime analyses were performed to examine the cell trajectory processes of neutrophils. In blood samples, AC111000.4, AKAP12, AL139042.1, BACE2, CA8, GATA2, GCSAML, HDC, S100A12, and S100A8 were identified as key genes involved in the fate determination of neutrophils in ICH patients (Fig. 5i-e). In hematoma samples, the top 10 key genes were AC111000.4, AKAP12, CAMK1D, GCSAML, HDC, MIF, MNDA, RIPOR2, SLC8A1 and SMAP2 during the fate determination of Neutrophils(Fig. 5ii-e). The shared gene, AKAP12 and HDC, have been reported to participated in the interaction with BBB [30, 31].
In blood samples, NK/T cells were re-clustered into four subtypes: CD4 + T cells (CD4), CD8 + T cells (CD8A, CD8B), Tregs (NCAM1), and NK cells (FOXP3) (Figure S6a). The top upregulated genes among these four subtypes were summarized, including LTB, LDHB, IL7R, GAS5, TCF7 for CD4 +; S100A8, S100A9, NAMPT, HBA2, CXCL8 for CD8 +; GNLY, GZMB, NKG7, FGFBP2, PRF1 for NK cells; and IL32, HLA-DRB1, LGALS3, ANXA2, TRAC for Tregs (Fig. 6a-i). This profile highlights biological functions beyond immune responses, including regulation of energy metabolism (LDHB) [32] and oxidative stress(NAMPT) [33]. Notably, CD8 + and NK cells exhibited higher proportions in ICH patients, while CD4 + cells were more prevalent in healthy controls (Fig. 6a-ii). For transcription factors, RCOR2, ZBED1, MYC, CIITA, and HIVEP2 were significantly upregulated in all four subtypes of NK/T cells, indicating regulation of the transcriptional steady state (RCOR2) [34], differentiation(ZBED1) [35] and metabolism(MYC) [36]. In contrast, RFXANK, RFX5, RFXAP, ING4, and KDM5A were upregulated only in Tregs, suggesting an immune suppressor role [37, 38] (Fig. 6a-iiii). GO and KEGG analyses revealed that DEGs of NK/T cells in ICH patients, compared with healthy controls, were primarily enriched in immune response processes, including regulating/activating signaling pathways and regulating cell surface receptor signaling pathways (Figure S6b-c). Additionally, biological functions such as ubiquitin ligase complex, ubiquitin-like protein transferase activity, and ubiquitin-protein transferase activity were activated, suggesting a potential key role of ubiquitin as a regulator of T cell differentiation and development in ICH [39, 40]. Ligand-receptor pairs of cell–cell interaction among NK/T cells in blood samples derived from ICH patients were shown in Figure S6d. In hematoma samples, NK/T cells were reclustered into three subtypes: CD4 + T cells (CD4), CD8 + T cells (CD8A, CD8B), and NK cells (FOXP3), based on marker genes (Figure S6e). In hematoma samples, the top five upregulated genes were TSHZ2, LEF1, LTB, IL7R, and CAMK4 in CD4 + T cells; CCL5, SGCD, CCL4L2, C1orf21, and HSPA1A in CD8 + T cells; and NKG7, GNLY, GZMB, PRF1, and KLRF1 in NK cells. The upregulation of CCL5 and CCL4L2 underscores their critical roles in mobilization and chemotaxis within CD8 + T cells (Fig. 6b-i). Among the three sub-clusters, CD8 + T cells exhibited a higher proportion in hematoma samples compared to blood samples (Fig. 6b-ii). Uniquely, CD8 + T cells exert direct cytotoxic effects and perforin-mediated neurotoxicity in focal lesions associated with cerebrovascular diseases [41, 42]. For transcription factors, RCOR2, MYC, ZBED1, TFAM, KLF13, CIITA, and HIVEP2 were upregulated across CD4 + T cells, CD8 + T cells, and NK cells (Fig. 6b-iii). Ligand-receptor pairs of cell–cell interaction among NK/T cells in hematoma samples derived from ICH patients were shown in Figure S6g.
Fig. 6
Compositional analysis of other immune cell types. Overview of NK/T cell subtypes. a(i) Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each NK/T cell subtypes in blood samples from ICH patients. (ii) comparison the proportion of NK/T cell subtypes from blood samples between ICH patients and healthy controls (iii) expression levels of transcript factors across NK/T cell subtypes in blood samples from ICH patients. b(i) Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each NK/T cell subtypes in hematoma. (ii) comparison the proportion of NK/T cell subtypes between blood and hematoma samples. (iii) expression levels of transcript factors across NK/T cell subtypes in hematoma samples from ICH patients. Overview of Monocytes/Macrophages subtypes. c(i) Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each Monocytes/Macrophages subtypes in blood samples from ICH patients. (ii) comparison the proportion of Monocytes/Macrophages subtypes from blood samples between ICH patients and healthy controls (iii) expression levels of transcript factors across Monocytes/Macrophages subtypes in blood samples from ICH patients. d(i) Volcano plot illustrating the top 5 up-regulated and down-regulated genes of each Monocytes/Macrophages subtypes in hematoma. (ii) comparison the proportion of Monocytes/Macrophages subtypes between blood and hematoma samples. (iii) expression levels of transcript factors across Monocytes/Macrophages subtypes in hematoma samples from ICH patients
In blood samples, Monocytes/Macrophages were categorized into classical and non-classical monocytes. In classical monocytes, VCAN, S100A8, S100A9, S100A12, and LYZ were identified as the top upregulated genes, while FCGR3A, CDKN1C, MS4A7, RHOC, and IFITM3 were the top upregulated genes in non-classical monocytes (Fig. 6c-i). For transcription factors, HIVEP2, MYC, and ZBED1 were upregulated in both classical and non-classical monocytes in ICH patients compared with healthy controls (Fig. 6c-iii). MAPK, chemokine, and AMPK signaling pathways were significantly activated in monocytes from ICH patients (Figure S6 i-j). Compared with the former two pathways, the AMPK signaling pathway is essential for restoring energy balance and reducing oxidative damage and inflammatory responses [43]. Similarly, ubiquitin-mediated proteolysis was also identified as an upregulated pathway.Ligand-receptor pairs of cell–cell interaction among Monocytes/Macrophages in blood samples derived from ICH patients were shown in Figure S6k. In hematoma samples, Monocytes/macrophages were categorized into classical, intermediate, and non-classical monocytes. For classical monocytes, the top upregulated genes included PLCB1, FKBP5, AOAH, RIPOR2, and LRRK2; for intermediate monocytes, SPP1, PPBP, CCL20, CXCL5, and CXCL8; and for non-classical monocytes, HLA − DPB1, SLC35F1, RPS2, RPS18, and RPS23. The RPS family plays a key role in protein synthesis and may also contribute significantly to the inflammatory response [44] (Fig. 6d-i). Among the three sub-clusters, classical monocytes displayed a higher proportion in hematoma samples compared to blood samples (Fig. 6d-ii).
Considering that Toll-like receptors 4(TLR4), which reported as an inflammatory marker in ICH by our team before [12], were reported to be associated with CXCL and Macrophage Migration Inhibitory Factor (MIF) signaling pathways [45, 46], cellular interaction of these two pathways were assessed both in blood and hematoma samples. For CXCL signaling pathways, Neutrophils have the higher expression than other cell types (Fig. 7a,d). The CXCL signaling pathways played key roles as sender, receiver and influencer in the interaction between Neutrophils and Monocytes or Platelets (Fig. 7c,f). The L-R pairs with the highest contribution in the CXCL signaling pathways were CXCL8-CXCR1/2 (Fig. 7b,e). CXCL family, a subtype of Chemokines, is known to participate in inflammatory process by regulating migrations of leukocytes including Neutrophils and Monocytes/Macrophages [47]. CXCL8, also known as IL-8, also plays roles as an angiogenic factor, as well as proinflammatory factor [48]. Besides, the release of CXCL8 depends on the expression of TLR4 during the process of inflammation [49]. Though the relationship between TLR4 and CXCL8 has rarely been reported in ICH, the significance of CXCL8 has confirmed as a predictor of neurological impairment severity and functional outcomes in ICH [50, 51], induced by heme oxygenase-1 released from hematoma [52].
Fig. 7
Cellular interactions of CXCL signaling pathway in ICH patients. Blood samples. a Outgoing and incoming signaling patterns for various cell types. b Contribution of each ligand-receptor pair. c Roles of CXCL signaling pathway network among various cell types. Hematoma samples. d Outgoing and incoming signaling patterns for various cell types. e Contribution of each ligand-receptor pair. f Roles of CXCL signaling pathway network among various cell types
For MIF signaling pathways, low cellular interactions were detected in blood samples from ICH patients (Fig. 8a,b). However, MIF signaling pathways showed high interaction in Neutrophils as sender while NK/T cells, Monocytes/Macrophages and B/Plasma cells played roles as receiver in hematoma samples (Fig. 8c,d,e). MIF, binding to CD74/CD44, is another key mediator of inflammation and oxidative stress in cerebrovascular disease [53]. The influence of MIF on neuron was divergent, both protective [54] and detrimental [55]. In hemorrhagic stroke, MIF levels are associated with inflammatory reaction, neurological injury and functional outcomes [56,57,58]. For Neutrophils, MIF-ACKR3 is also essential in inflammatory process [59]. For MIF, TLR4 was also reported as a key regulator in the process of inflammation with different immune cells including T cells [60], microglial [55] and Monocytes/Macrophages [
Comments (0)