
Remember me
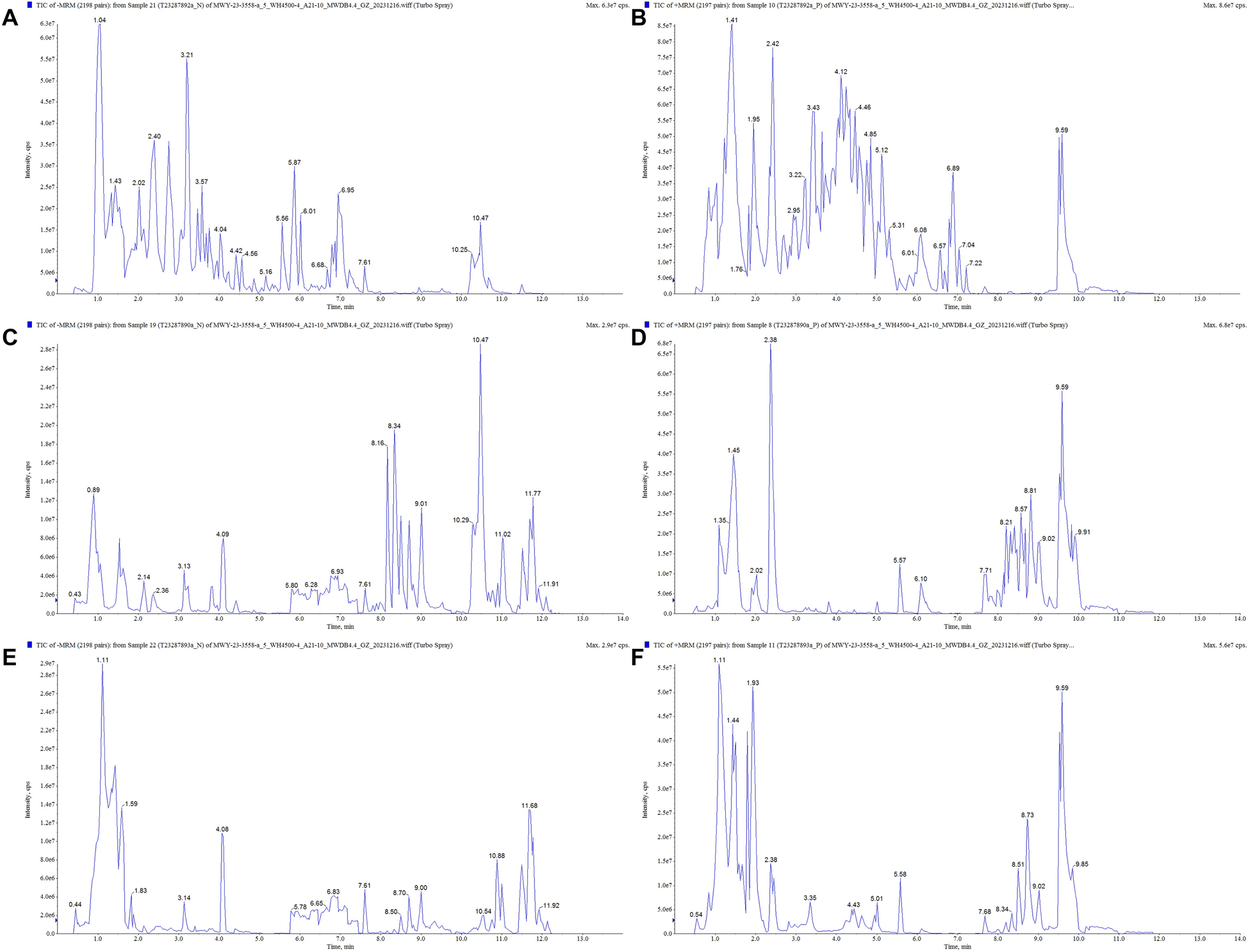
In order to deeply explore the material basis of the anti-AD efficacy of HLJDD, we performed UPLC-MS/MS analysis with gradient elution using 0.1% formic acid aqueous solution-0.1% formic acid acetonitrile solution as the mobile phases, and full scans of the extract of HLJDD, brain tissue of APP/PS1 mice, and plasma of APP/PS1 mice were carried out in both the positive and negative ionization modes, and the sample base-peak ion the total ion current (TIC) chromatograms are shown in Fig. 1A–F. A total of 137 chemical constituents were identified from HLJDD (Table. S1), including 51 flavonoids, 22 terpenoids, 20 alkaloids, 24 organic/phenolic acids, 6 lignans/coumarins, and 14 other compounds. Of these, 49 components were found in the brain tissue of APP/PS1 mice (Tables S2), 48 components were found in the plasma of APP/PS1 mice (Table S3). And 42 of these compound components were simultaneously found in the blood and brain tissues (Table S4).
Fig. 1
Chemical composition of HLJDD was determined. A TIC diagram of negative ion mode of HLJDD. B TIC diagram of positive ion mode of HLJDD. C TIC diagram of drug-containing serum negative ion pattern. D TIC diagram of drug-containing serum positive ion pattern. E TIC map of negative ion pattern in brain tissue. F TIC map of positive ion patterns in brain tissue
Network pharmacology study of HLJDD on APP/PS1 miceTo preliminarily explore the mechanism of action of HLJDD in the treatment of AD, we employed network pharmacology to predict potential pathways of action (Fig. 2A). A total of 36 components that can enter the bloodstream and brain were selected, excluding six components that lacked potential targets. We identified 507 targets associated with these components and 15,318 targets related to AD, resulting in the identification of 466 overlapping targets (Fig. 2B). The component-target-pathway map was constructed using Cytoscape 3.9.1 software. In Fig. 2C, circles represent the components, rhombuses denote the related pathways, squares indicate the intersection targets, triangles represent HLJDD, and inverted triangles signify AD. We predict that the core components with significant therapeutic potential include: (12bs)−4,10,11-trimethoxy-7,8,12B,13-tetrahydro-5 h-6-azatetraphen-3-ol, Baicalein, Epiberberine, 3’,7-dihydroxy-4’-methoxyflavone, Thaliporphine, Wogonin, Kumatakenin, Jatrorrhizine, 6-ethyl-1,10-dimethoxy-5,6,6A,7-tetrahydro-4H-dibenzo[de,g]quinoline-2,9-diol, phellodendrine, and Hamiltone A. The 466 overlapping targets of HLJDD and AD were input into the STRING database to generate a protein–protein interaction (PPI) graph comprising 92 nodes and 1,947 edges (Fig. 3D). The key intervention targets primarily include GAPDH, AKT1, TNF, SRC, EGFR, BCL2, STAT3, HSP90AA1, CASP3, and JUN, et al. (Table S5).
Fig. 2
Study on network pharmacology of HLJDD against APP/PS1 mice. A Images of the various medicinal materials of HLJDD. B Venn plot of the intersection target of HLJDD and AD, where the green circles represent the active ingredient targets, and the blue circles represent the targets related to AD. C Image of the blood-entry and brain-entry component-target—disease pathway of HLJDD. The circles represent the blood-entry components, the rhombus represents the related pathways, the square represents the intersection targets, the triangle represents HLJDD, and the inverted triangle represents AD. D PPI network diagram of the core target. E Analysis diagram of the potential target GO pathway of HLJDD in the treatment of AD. F Bubble plot of KEGG pathway analysis for potential targets of HLJDD in the treatment of AD
Fig. 3
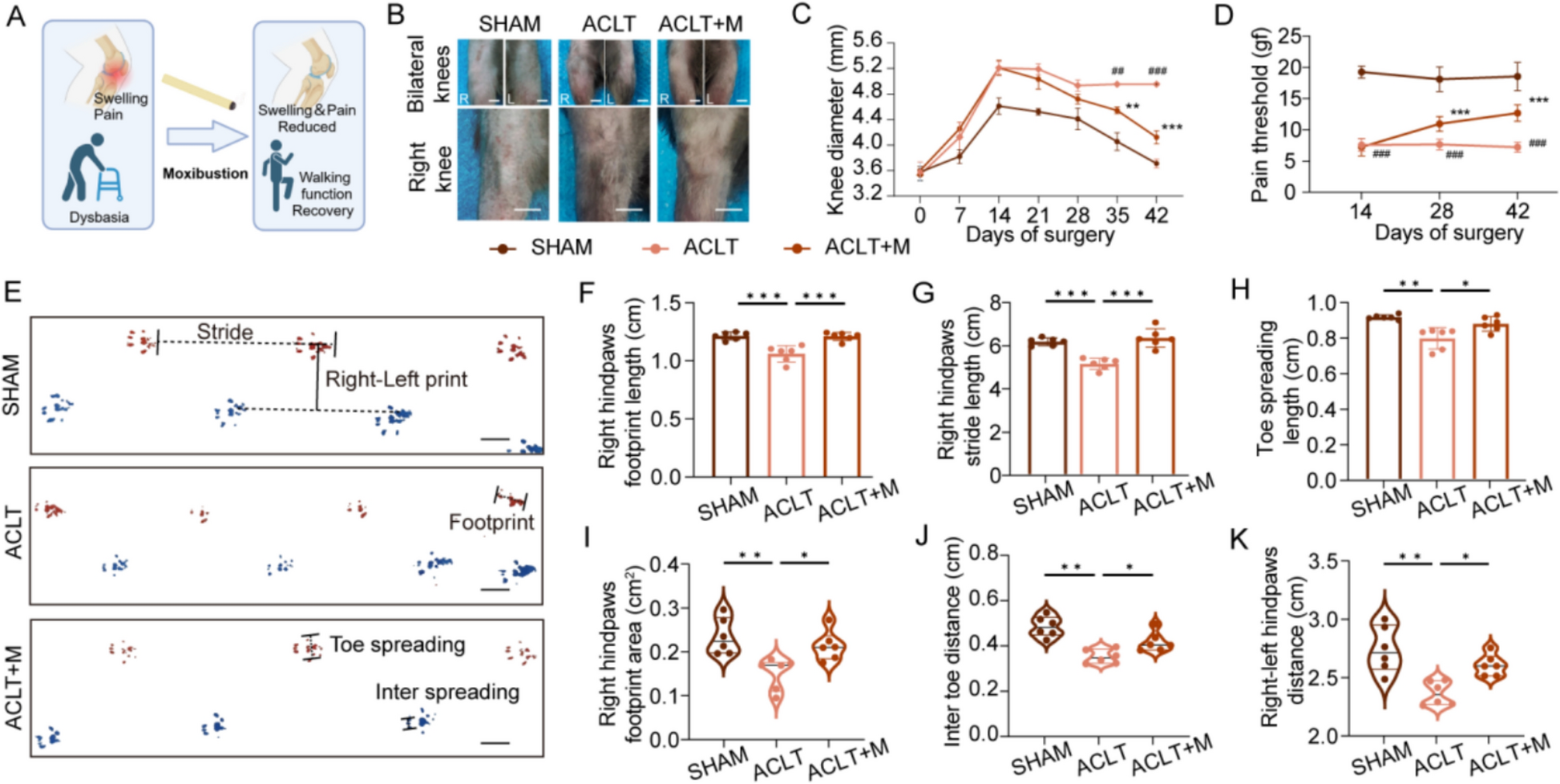
Effects of HLJDD on cognitive function in APP/PS1 mice. A Morris water maze was used to detect behavioral changes in mice. B Escape latency analysis of mice in each group. C The swimming trajectory of the mice on the 6th day within 60 s. D Data analysis of the time spent in the target quadrant on the sixth day. E The number of platform crossings in mice on the sixth day. F Representative images of the nesting experiment of mice from 0 to 3 days. G The quantitative analysis results of the nesting score. Data are presented as mean ± SD and data are presented as mean ± SD (n = 6). ##P < 0.01 (vs control group), *P < 0.05 and **P < 0.01 (vs model group)
In the GO functional enrichment analysis, a total of 1,031 biological processes (BPs) may be influenced by HLJDD. These processes primarily include protein phosphorylation, response to xenobiotic stimuli, insulin-like growth factor receptor signaling pathways, positive regulation of the MAPK cascade, epidermal growth factor receptor signaling pathways, and G protein-coupled receptor signaling pathways coupled to cyclic nucleotide second messengers. Additionally, there are 137 cellular components (CCs) identified, which mainly encompass the plasma membrane, dendrites, cytosol, perinuclear region of the cytoplasm, synapses, and receptor complexes. The Molecular Functions (MF) category consists of 354 items, including histone H3Y41 kinase activity, histone H2AXY142 kinase activity, protein kinase activity, protein tyrosine kinase activity, protein serine kinase activity, and ATP binding. The results of the KEGG pathway enrichment analysis indicated that there are a total of 181 pathways relevant to treating AD with HLJDD. The enrichment analysis results of the top 20 KEGG pathways were input into microbiome bioinformatics data analysis platform, generating a bubble chart for KEGG pathway enrichment analysis. From Fig. 2F, it is evident that the primary signaling pathways involved in the treatment of AD by HLJDD include pathways in cancer, lipid and atherosclerosis, neuroactive ligand-receptor interactions, serotonergic synapses, the PI3K-Akt signaling pathway, the Ras signaling pathway, and the MAPK signaling pathway.
Effects of HLJDD on cognitive function in APP/PS1 miceIn the present study, the effects of HLJDD on learning memory ability of AD model mice were evaluated by Morris water maze experiment. The results showed that compared with the blank group, mice in the model group and the HLJD-L exhibited significant spatial cognitive deficits manifested by disturbed motor trajectories (Fig. 3A), prolonged avoidance latency (Fig. 3B), as well as shortened residence time in the target quadrant and reduced number of traversing platforms (Fig. 3C–E). On the other hand, mice in the positive drug group, HLJD-M and HLJD-H showed significant improvement in cognitive function, and their behavioral performance was similar to that of the control group, which was characterized by a more concise motor trajectory, significantly shorter evasion latency, and the ability to maintain the memory of the original platform position after the platform was removed. These results suggest that the spatial learning and memory ability of APP/PS1 mice can be effectively improved by HLJD-M and HLJD-H.
We further used the nesting experiment to assess the cognitive function of AD mice. The experimental results showed that during the 24-h observation period, the mice in each group showed only a small amount of action. At 48 h, mice in the control group, the positive drug group, HLJD-H and HLJD-M were able to complete the initial construction of their nests and maintain their structural integrity, and at 72 h, mice in the control group, the positive drug group, the HLJD-M group and the HLJD-H group successfully constructed well-constructed nests (Fig. 3F), and the results of the nest-building scores were in line with the observed phenomena (Fig. 3G). The combination of the results of the water maze and nest building experiments confirmed that HLJDD treatments significantly improved the cognitive dysfunction of the AD model mice, allowing them to regain near-normal levels of cognitive ability after a short period of training. This finding further supports the ameliorative effect of HLJDD on cognitive impairment in AD.
Effects of HLJDD on hippocampal neural structure and Aβ deposition in APP/PS1 miceHistopathological analysis revealed the mechanism of action of HLJDD on neuroprotection in AD model mice. HE staining results showed that the cortex, hippocampus and CA1 area of the model group mice presented typical features of neuronal damage, which was manifested as cellular crumpling and sparse arrangement. After HLJDD intervention, neuronal morphology and distribution were significantly improved (Fig. 4A). The results of Nissl staining showed that HLJDD had no significant promoting effect on hippocampal neurons and hippocampal neurogenesis (Fig. 4B). Immunohistochemical analysis showed that HLJDD significantly reduced Aβ plaque deposition in the whole brain and hippocampus of APP/PS1 mice, with the most significant improvement in the HLJD-M and HLJD-H groups (Fig. 4C). These results suggest that HLJDD may exert neuroprotective effects through multiple pathways such as attenuating neuronal damage and inhibiting Aβ deposition, thereby improving AD-related pathological features.
Fig. 4
Neuroprotective effect of HLJDD on APP/PS1 mice. A Representative pictures of HE staining of brain tissue in mice, scale bar = 100 μm (n = 6). B Representative pictures of Nissl staining of brain tissue in mice, scale bar = 100 μm (n = 6). C Representative pictures of Aβ deposition of brain tissue in mice, scale bar = 500 μm (n = 6)
Effects of HLJDD on central neuroinflammation in APP/PS1 miceAstrocytes and microglia are crucial immune cells in the central nervous system, playing a pivotal role in the regulation of neuroinflammation. To investigate the effects of HLJDD on these two cell types, we employed immunofluorescence staining techniques to assess its regulatory effects on the expression of the astrocyte marker GFAP and the microglial marker Iba-1. The experimental results demonstrated (as depicted in Fig. 5A, B) that the expression levels of GFAP and Iba-1 in the hippocampus of mice in the model group were significantly elevated, indicating an active state of neuroinflammation. Following HLJDD intervention, the expression levels of Iba-1 and GFAP in the hippocampus of mice were significantly reduced, suggesting that HLJDD has a notable inhibitory effect on the activation of microglia and astrocytes. Furthermore, the mechanisms by which astrocytes and microglia contribute to neuroinflammation in Alzheimer’s disease are closely related to their subtypes. Therefore, we further analyzed the expression of different cell subtypes in the hippocampus of mice post-HLJDD intervention. Specifically, we examined the expression levels of type A1 astrocytes (GFAP+/C3+), type A2 astrocytes (GFAP+/S100A10+), type M1 microglia (Iba-1+/iNOS+), and type M2 microglia (Iba-1+/Arg-1+). The results indicated (as shown in Fig. 5A, B) that HLJDD could significantly inhibit the expression of GFAP+/C3+ and Iba-1+/iNOS+, while promoting the expression of GFAP+/S100A10 and Iba-1+/Arg-1+. This finding suggests that HLJDD may exert its anti-neuroinflammatory effects by modulating the subtype balance of astrocytes and microglia.
Fig. 5
Effects of HLJDD on central neuroinflammation in APP/PS1 mice. A Expression of GFAP/C3 and GFAP/S100A10 in hippocampus of mice in each group (The white arrow indicates double positive staining). B Expression of Iba-1/Arg-1 and Iba-1/iNOS in hippocampus of mice in each group (The white arrow indicates double positive staining). C The infiltration of CD4+T and CD8+T cells in hippocampus of mice in each group. D Levels of IL-1β, IL-6, IL-4, and IFN-γ in brain tissue. Data are presented as mean ± SD (n = 6). scale bar = 100 μm. ##P < 0.01 and ###P < 0.001 (vs control group), *P < 0.05, **P < 0.01 and ***P < 0.001 (vs model group)
The permeability of BBB in patients with AD is elevated, allowing peripheral intrinsic and adaptive immune cells to infiltrate the brain. This infiltration of peripheral immune cells contributes to and mediates neuroinflammation, thereby significantly influencing the progression of AD. In this study, we observed marked infiltration of CD4+ and CD8+ T cells in the brains of mice in the model group. However, following intervention with HLJDD, the infiltration of CD4+ and CD8+ T cells in the mouse brains was significantly reduced (as illustrated in Fig. 5C). Additionally, we conducted a quantitative analysis of the expression levels of inflammatory factors in brain tissue, including pro-inflammatory factors IL-1β, IL-6, IFN-γ, and the anti-inflammatory factor IL-4. The results are presented in Fig. 5D. Compared to the model group, the HLJDD intervention led to a significant decrease in the expression levels of pro-inflammatory factors in the mouse brain, while the expression levels of anti-inflammatory factors increased, resulting in a notable reversal of the overall inflammatory state (P < 0.05). These findings suggest that HLJDD can effectively mitigate central nervous system inflammation in AD mice.
Effects of HLJDD on peripheral inflammation in APP/PS1 miceWe further evaluated the improvement of peripheral inflammation following HLJDD intervention by analyzing the number of immune cells in the peripheral blood and the expression levels of inflammatory factors in the serum of mice across all groups. Our findings indicate that, compared to the blank group, the model group exhibited a significant increase in the number of immune cells in peripheral blood (Fig. 6A). Additionally, the expression levels of pro-inflammatory factors IL-1β, IL-6, and IFN-γ in serum were markedly elevated, while the expression level of the anti-inflammatory factor IL-4 was significantly reduced (Fig. 6B). This suggests an abnormal peripheral inflammatory response in AD mice. Following HLJDD intervention, a comparison with the model group revealed that the numbers of peripheral blood white blood cells, lymphocytes, monocytes, and granulocytes in the various HLJDD dose groups all decreased (Fig. 6A). Furthermore, the expression levels of peripheral blood inflammatory factors IL-1β, IL-6, IL-4, and IFN-γ in the HLJDD dose groups were all reversed compared to those in the model group (Fig. 6B). These results suggest that HLJDD exerts a significant inhibitory effect on the level of peripheral inflammation in AD mice.
Fig. 6
Effects of HLJDD on peripheral neuroinflammation in APP/PS1 mice. A Levels of IL-1β, IL-6, IL-4, and IFN-γ in peripheral serum. B Blood routine was used to detect the number of immune cells. Data are presented as mean ± SD (n = 6). #P < 0.05, ###P < 0.001 and ####P < 0.0001 (vs control group), *P < 0.05 and **P < 0.01 (vs model group)
Effects of HLJDD on the intestinal barrier of APP/PS1 miceAs shown in Fig. 7A, the structure of the small intestinal epithelium in the model group of mice was significantly disorganized and atrophied, however, the intestinal epithelial villi damage was improved by treatment with different concentrations of HLJDD. Immunohistochemical results showed that the expression of Occludin and ZO-1 was reduced in the model group compared with the control group, and improved after treatment. The results of western blot showed that the expression of Occludin and ZO-1 expression was downregulated in the model group compared to the control group, whereas the protein expression was upregulated after treatment (Fig. 7B–D, P < 0.05).
Fig. 7
Effect of Huanglian-Jiedu decoction on intestinal barrier in APP/PS1 mice. A Representative image of HE staining in the mouse intestine, and representative images of immunohistochemistry of intestinal Occludin and ZO-1 (Scale bar = 50 μm). B Statistical analysis of the relative expression level of Occludin. C Statistical analysis of the relative expression level of ZO-1. D The expressions of ZO-1 and Occludin proteins in the intestine were detected by Western blotting. E Semi-quantitative analysis of the gray value of Occludin. F Semi-quantitative analysis of the gray value of ZO-1. Data are presented as mean ± SD (n = 3). ##P < 0.01 (vs control group), *P < 0.05 and **P < 0.01 (vs model group)
Effects of HLJDD on gut microbiota in APP/PS1 miceThe results of alpha and beta diversity analyses based on 16S rRNA gene sequencing showed that HLJDD had a significant effect on the composition of the gut microbiota. In terms of alpha diversity, changes in Chao1 index and Shannon index indicated that the model group varied from the treatment group in terms of the number of operational taxonomic units (OTUs) as well as the abundance and diversity of species (Fig. 8A, B). Beta diversity analyses further revealed that there were significant clustering differences in microbial taxonomic and functionality among the different groups through the PCOA three-dimensional maps (Fig. 8C). To estimate the distribution of mouse gut microbes in each group, we compared the relative abundance of the top 30 groups at the phylum and genus levels (Fig. 8D–G). At the phylum level, the abundance of the Firmicutes group in the model group was significantly reduced compared with the control group (P < 0.01), while the treatment group brought back its abundance, on the contrary, the abundance of the Rokubacteria group in the model group was significantly elevated (P < 0.05), and was treated and significantly decreased (Fig. 8H, I, P < 0.05). At the genus level, the abundance of Bacteroides was elevated but not statistically significant in the treatment group compared to the model group, however, the abundance of Lachnospiraceae decreased in the model group showed an upward trend after treatment (Fig. 8J, K). Taken together, these results suggest that HLJDD was able to modulate the structure of the gut microbiota and restore part of the disturbed microbial composition in the model group.
Fig. 8
Effects of HLJDD on gut microbiota in APP/PS1 mice. A Chao 1 index. B Shannon index. C The principal co-ordinates analysis (PCoA) plot. D and E Heatmaps and histograms of gut microbiota composition at the phylum level (top 30 relative abundance). F and G Heatmaps and histograms gut microbiota composition at the genus level (top 30 relative abundance). H–K Relative abundance of Firmicutes, Rokubacteria, Bacteroides and Lachnospiraceae. Data are presented as mean ± SD (n = 6). *P < 0.05, **P < 0.01 and ***P < 0.001
Effects of HLJDD on the expression of inflammation-related proteins in APP/PS1 miceTo further explore the interaction between HLJDD, NLRP3, and Caspase-1, we conducted a molecular docking analysis. Based on the degree value index, we screened the top-ranked active ingredients in HLJDD as core components for treating AD. The active ingredients include (12bs)−4,10,11-trimethoxy-7,8,12B,13-tetrahydro-5 h-6-azatetraphen-3-ol, Baicalein, Epiberberine, 3’,7-dihydroxy-4’-methoxyflavone, Thaliporphine, Wogonin, Kumatakenin, Jatrorrhizine, 6-ethyl-1,10-dimethoxy-5,6,6a,7-tetrahydro- 4H-dibenzo[de,g]quinolin-2,9-diol, phellodendrine, and Hamiltone A, which were molecularly docked with NLRP3 and Caspase-1, respectively. A binding energy threshold of −5 kcal/mol was considered indicative of good binding activity. The heat map of the docking results is shown in Fig. 9A. It was found that the binding energies of the compounds with NLRP3 were all lower than −5 kcal/mol, indicating that the main active components of HLJDD exhibit significant binding activity with NLRP3. Subsequently, the three complexes with the highest docking scores and binding energies were selected for further visualization (Fig. 9B–D). Next, we validated the expression of inflammation-related proteins in the intestines using Western blot (Fig. 9E). The expression of NLRP3, Caspase-1 and ASC proteins was significantly up-regulated in the model group compared to the control group, whereas HLJDD intervention significantly reversed these changes (Fig. 9F).
Fig. 9
Effect of HLJDD on the expression of inflammation-related proteins in APP/PS1 mice. A Heat maps of the binding energies of each chemical component with NLRP3 and Caspase-1. B–D 3D and 2D interaction diagrams of the core compound and NLRP3 protein, including, B Jatrorrhizine, C Kumatakenin, D 3’,7-dihydroxy-4’-methoxyflavone. E The protein expression of NLRP3, Caspase-1 and ASC in the intestines was determined by Western blotting (n = 3). F grey-scale quantitative analysis of index. Data are presented as mean ± SD. #P < 0.05 and ###P < 0.001 (vs control group), *P < 0.05 (vs model group)
Comments (0)