
Remember me
As one of the important organs of the human body, the kidney is crucial in regulating the balance of water and electricity, filtering wastes, and participating in metabolic processes. Renal cell carcinoma (RCC) affects the urinary system and has three primary subtypes, of which clear cell RCC represents the majority (approximately 75%).[1,2] The incidence of RCC has increased recently. RCC affects nearly 300,000 people worldwide and causes more than 100,000 deaths each year.[3] The mutant genes in RCC are related to cell metabolism and participate in iron, oxygen, energy, and nutrient metabolism, involving glycolysis, tricarboxylic acid cycle, and other processes, which are called metabolic diseases and are the model for studying the Warburg effect.[4,5] Cancer cells usually require a large amount of substances and energy to maintain their need for rapid proliferation. In RCC, the genes of the tricarboxylic acid cycle pathway are downregulated, and mitochondrial oxidative phosphorylation is impaired.[2,6] Cancer cells supply energy and substances through the pentose phosphate pathway to promote the growth of tumor cells.[7] In renal cancer, glycolysis-related genes are upregulated, and the glycolysis and lactate production of renal cancer cells and the utilization rate of glucose increase.[8] Therefore, aerobic glycolysis-targeted therapy for renal cancer is an effective clinical treatment for tumors.
Phosphoinositol-3-kinase (PI3K)/protein kinase B (AKT)/ mammalian target of rapamycin (mTOR) pathway participates in various physiological activities, such as proliferation, metabolism, and metastasis in cancer.[9] This signaling pathway is often overactivated in renal cancer,[10] and blocking it can restrain the malignant behavior of RCC and induce autophagy and apoptosis of human RCC cells.[11] Wang et al.,[12] found that glycolysis, facilitated by the PI3K/ AKT/mTOR cascade, accelerates the progression of lung cancer. Other studies found that DEPDC1 might influence glycolysis through the AKT/mTOR/HIF1α pathway.[13] These findings suggest a connection between this signaling axis and glycolysis regulation in RCC.
Ubiquitin C-terminal hydrolase L5 (UCHL5), one of the four members of the UCHL family, is present in the cytoplasm and nucleus and has a significant function in tumorigenesis and metastasis.[14-17] Evidence reveals that UCHL5 is implicated in multiple cancer types, including bladder, endometrial, lung, stomach, and colon cancers. For instance, UCHL5 affects the biological behavior of cervical cancer cells and apoptosis.[18] Elevated UCHL5 levels are associated with poor prognosis in patients with lung adenocarcinoma (LUAD), and silencing UCHL5 markedly inhibits LUAD cell growth.[19] UCHL5 is involved in glycolysis and affects the development of hepatocellular carcinoma.[20] In bladder cancer, UCHL5 promotes the growth and metastasis through the AKT/mTOR signaling pathway.[21] Zhang et al., found that UCHL5 is upregulated in patients with advanced RCC, so it may be a prognostic marker for RCC.[14] Nevertheless, limited research has been conducted on the mechanisms of UCHL5 in RCC. This study aims to investigate the expression of UCHL5 in RCC and the involved biological processes to provide new insights for developing new treatment methods.
MATERIAL AND METHODS Cell cultureIn brief, RCC cell lines (786-O, iCell-h235) and renal tubular epithelial cell lines (Hexokinase-2 [HK-2], iCell-h096) were purchased from Cellverse Co., Ltd. (Shanghai, China). The 786-O cells were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (21875042, Gibco, USA), and HK-2 cells were cultured in Dulbecco’s Modified Eagle Medium (11995065, Gibco, USA). The medium contained 10% fetal bovine serum ([FBS], 30067334, Gibco, USA) and penicillin– streptomycin (P1400, Solarbio, Beijing, China) at 37°C and 5% CO2. All cells were validated for authenticity by short tandem repeat and were free of mycoplasma contamination.
Real-time quantitative polymerase chain reaction (qRT-PCR)First, RNA was extracted using TRIzol® reagent (15596026, Thermo Fisher Scientific, USA). Next, complementary DNA generation and qRT-PCR were performed using One Step PrimeScript™ RT-PCR Kit in accordance with the manufacturer’s oper ation manual (RR064A, Takara, Japan). The primers used were as follows: Forward 5’-CAGTGCAGTAAGGCCTGTCA-3’, and Reverse 5’-CCATGGGTTCCTCTGCAAGT-3’ for UCHL5; Forward 5’-GTGGACATCCGCAAAGAC-3’, and Reverse 5’-GAAAGGGTGTAACGCAACTA-3’ for β-actin. Relative messenger RNA (mRNA) expression was assessed by the 2-ΔΔCt calculation with β-actin as reference.
Western blot (WB) analysisExperimental procedures were performed as described previously.[22] Protein was extracted, and its concentration was detected using radio-immunoprecipitation assay ( RIPA) lysis buffer (P0013B, Beyotime, Shanghai, China) and bicinchoninic acid ( BCA) method (P0010S, Beyotime, Shanghai, China). The proteins were subjected to 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA) using the wet transfer method. The membranes were incubated with 5% skimmed milk for 60 min, and the specific primary antibodies were incubated at 4°C overnight. The following specific primary antibodies were obtained from Proteintech (Wuhan, China) at a dilution ratio of 1:1000 as required by the manufacturer: Anti-UCHL5 (11527-1-AP), anti-glucose transporter 1 (GLUT1) (81463-1-RR), anti-HK2 (22029-1-AP), anti-pyruvate kinase M2 (PKM2) (15822-1-AP), anti-lactate dehydrogenase A (LDHA) (84198-3-RR), anti-AKT (10176-2-AP), anti-p-AKT (80455-1-RR), anti-mTOR (81670-1-RR), anti-p-mTOR (28879-1-AP), anti-PI3K (67071-1-lg), anti-p-PI3K (HA721672), and anti-β-actin (66009-1-lg). The membranes were incubated with the corresponding horseradish peroxidase (HRP)-conjugated secondary antibodies (goat anti-mouse immunoglobulin G [IgG] H&L [HRP] [ab205719, 1:5000, Abcam] and goat anti-rabbit IgG H&L [HRP] [ab205718, 1:5000, Abcam]) at 37°C for 30 min. The protein bands on the membrane were detected with enhanced chemiluminescence luminescent reagent (WBKLS0500-2, Millipore, USA) and captured using the Tanon-4600 imaging system (Tanon-4600, Tanon, Shanghai, China). ImageJ software (National Institutes of Health, Bethesda, Maryland, USA) was used for quantitative gray value measurement, with β-actin serving as the internal reference. to calculate the relative expression of the target protein.
Overexpression and knockdown of UCHL5UCHL5 overexpression and knockdown lentivirus were acquired from Sangon Biotech Co., Ltd. (Shanghai, China). In brief, 786-O cells were transfected with lentiviral vectors; the transfection protocol was carried out as recommended by the manufacturer (R00430, Yunzhou biological, Guangzhou, China). The transfection sequence has been added to the Supplementary Materials. Effective integration of lentivirus transfection was detected by qRT-PCR and WB to verify the efficacy of the UCHL5 lentivirus transfection.
Colony formation assayThe 786-O cells were prepared into 90 cells/mL, and 1 mL from each group was inoculated and cultured in 24-well plates. After 21 days, the medium was discarded, rinsed, and fixed, and crystal purple (C0121, Beyotime, Beijing, China) staining was performed. The cloning formation of cells was imaged using a microscope (CX23, Olympus, Tokyo, Japan), and the number of clones formed was quantified by Image J.
5-ethynyl-2’-deoxyuridine (EdU) assayThe suspended cells were inoculated into 96-well plates and incubated at 37°C overnight. The cells were added with EdU reagent (C0081, Beyotime, Beijing, China) and incubated again for 90 min. The cells were then incubated with 4% paraformaldehyde (P0099, Beyotime, Shanghai, China) and permeabilization buffer (P0097, Beyotime, Beijing, China) for 15 min and then washed with phosphate-buffered saline (PBS). After being washed with PBS, EdU staining was performed. 4’,6-diamidino-2-phenylindole, dihydrochloride ( DAPI) solution (D1306, Invitrogen, Carlsbad, CA, USA) was added into the cells to stain the nuclei. Cell proliferation was observed by fluorescence microscopy (CKX53, Olympus, Tokyo, Japan).
Transwell assayThe Transwell chamber (3401, Corning, NY, USA) was placed in the 24-well plate, and the upper chamber was precoated with 500 ng/µL Matrigel (354480, Corning, NY, USA). The experimental procedures are based on previous literature.[23] In brief, 5 × 105 cells/mL cell suspensions were prepared and inoculated in the upper chambers, and the lower chambers were added with the medium containing 10% FBS. After a 24-h incubation, the cells were rinsed, fixed, and stained with crystal purple. Observations and photography were conducted under a microscope (CX23, Olympus, Tokyo, Japan) for subsequent statistical evaluation.
Cell metabolism assayGlucose and lactate concentrations were assessed to analyze alterations in cellular glucose metabolism. The cell supernatant of each treatment group was collected, and glucose and lactate levels were quantified using commercial test kits (A019-2-1 and A154-1-1) according to the manufacturers’ protocols (Nanjing Jiancheng Institute, Nanjing, China). Optical density was recorded using a microplate reader (VLBLATGD2, Thermo Fisher Scientific, MA, USA).
Tumor formation in nude miceBALB/c nude mice (male, 4–6 weeks of age, 18–22 g) were cultured in the light/dark cycle of 12/12 h (45% ± 5% humidity and 20 ± 2°C temperature). Mice were randomly assigned into two groups (n = 6/group) and administered with subcutaneous injection in the back with either 786-O cells stably expressing sh-UCHL5 or with negative control cells (1 × 107 cells per mouse). Mice were sacrificed by intraperitoneal injection of pentobarbital sodium (P3761, Sigma, Merck, USA) at a dose of 110 mg/kg according to their body weight to isolate tumors. Tumor dimensions were measured, and volume was calculated using the formula: (length × width2)/2. After 4 weeks, all mice were euthanized for tumor extraction and further analyses. All animal protocols were conducted under institutional ethical standards approved by The Experimental Animal Welfare Ethics Committee of The First Hospital Affiliated with Shandong First Medical University (Approval No. 2024111302).
Immunohistochemistry (IHC)The isolated transplanted tumor tissue was fixed with 4% paraformaldehyde and prepared into paraffin sections. After dewaxing and hydration, the sections were sequenced for antigen repair, incubated with 3% hydrogen peroxide solution for 10 min, and blocked with 2% BSA for 30 min at 37°C. The primary antibodies were purchased from HUABIO company and included anti-UCHL5 (HA721077), anti-GLUT1 (ET1601-10), anti-HK2 (ER1803-23), anti-PKM2 (ER1802-70), and anti-LDHA (ER00702) and incubated overnight at 4°C. After washing with Tween-20 phosphate-buffered saline 3 times, the section was incubated with secondary antibodies (GB23303, Servicebio, Wuhan, China) at room temperature for 10 min. The section was then added with 3,3’-Diaminobenzidine (P0203, Beyotime, Shanghai, China) for development, observation, and photography under microscope. Immunohistochemical staining was evaluated using the following criteria: The staining intensity was scored from 0 to 3, with no color (0), light yellow (1), brown (2), and dark brown (3). The proportion of positive cells within the same field was graded on a scale of 1–4. Specifically, cell percentages of <5%, 5–25%, 25–50%, 50–75%, and >75% were given scores of 0, 1, 2, 3, and 4, respectively. The two individual scores were multiplied to determine the visual field score. The overall score per field was determined by multiplying intensity and proportion scores. The final pathological score was obtained by averaging the results from five randomly selected fields.
Statistical analysesGraphPad Prism 9.0 software (San Diego, CA, USA) was utilized for statistical evaluation, and results were expressed as mean ± standard deviation. The t-test was used to compare two groups, whereas one-way analysis of variance, followed by Tukey’s post hoc test, was used to compare multiple groups. A P-value below 0.05 was deemed to reflect a statistically significant difference.
RESULTS UCHL5 expression was upregulated in RCC cellsTo explore the expression pattern of UCHL5 in RCC, we selected 786-O and HK-2 cells and detected differences in UCHL5 expression at protein and mRNA levels, respectively. The qRT-PCR and WB results showed the high expression of UCHL5 in 786-O cells than in HK-2 cells (P < 0.01 [Figure 1]).

Export to PPT
Knockdown of UCHL5 inhibited the malignant behavior of RCC cellsTo explore the role of UCHL5 in RCC, we examined how altering UCHL5 expression through gene overexpression and knockdown affects the malignant behavior of RCC cells. The WB and qRT-PCR data verified the efficiency of overexpression or knockdown in 786-O cells. No obvious differences were observed between the control group and either the overexpression negative control ( OE-NC) or short hairpin RNA negative control ( sh-NC) groups [Figure 2a-f]. UCHL5 expression markedly increased in the OE-UCHL5 group than in the OE-NC group and significantly reduced in the sh-UCHL5 group relative to sh-NC (P < 0.001). In particular, sh-UCHL5-2 was selected for the following experiments because it had the highest transfection (P < 0.001). Colony formation assays revealed that elevating UCHL5 expression significantly promoted colony formation, whereas its knockdown notably diminished this ability (P < 0.01 [Figure 2g and h]). The EdU assay showed that the proliferation of 786-O cells was accelerated after overexpression of UCHL5, while the proliferation of 786-O cells in the sh-UCHL5 group was slowed down (P < 0.05, [Figure 2i and j]). The Transwell assays demonstrated reduced invasive cells in the sh-UCHL5 group, whereas UCHL5 overexpression led to increased invasion relative to the OENC group (P < 0.001) [Figure 2k and l].
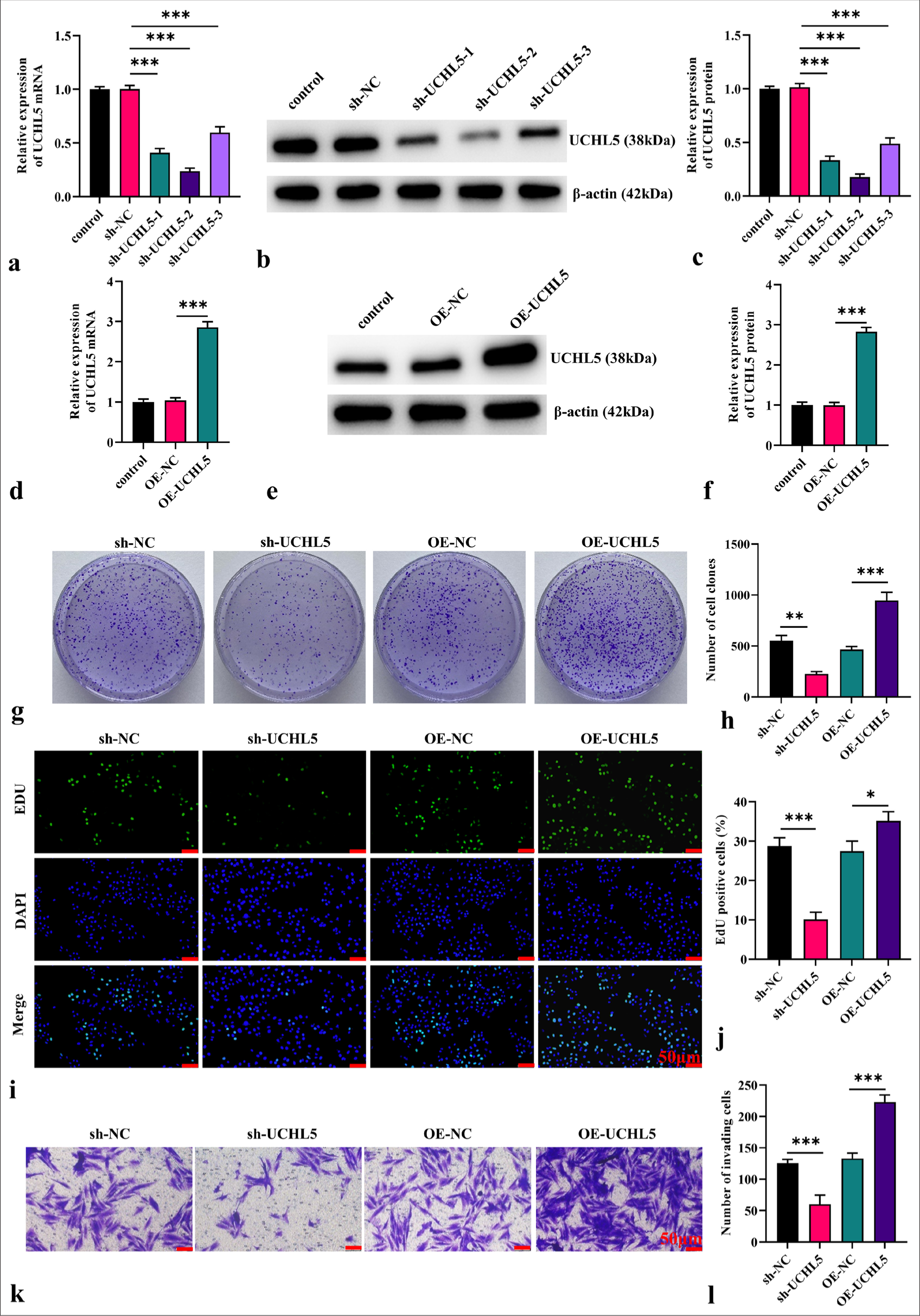
Export to PPT
Knockdown of UCHL5 inhibited the glycolysis of RCC cellsTo explore the correlation between UCHL5 and glycolysis in RCC cells, we first measured glucose and lactate levels in the cell culture system. UCHL5 overexpression increased glucose consumption and lactate production (P < 0.001), whereas knocking down UCHL5 produced the opposite result (P < 0.001 [Figure 3a and b]). The WB analysis results demonstrated the reduced levels of glycolysis-related protein in the sh-UCHL5 group (P < 0.01). By contrast, the OE-UCHL5 group had elevated expressions of glycolysis-related proteins (P < 0.01 [Figure 3c and d]). This finding suggests a positive correlation between UCHL5 expression and glycolysis activity in RCC cells.
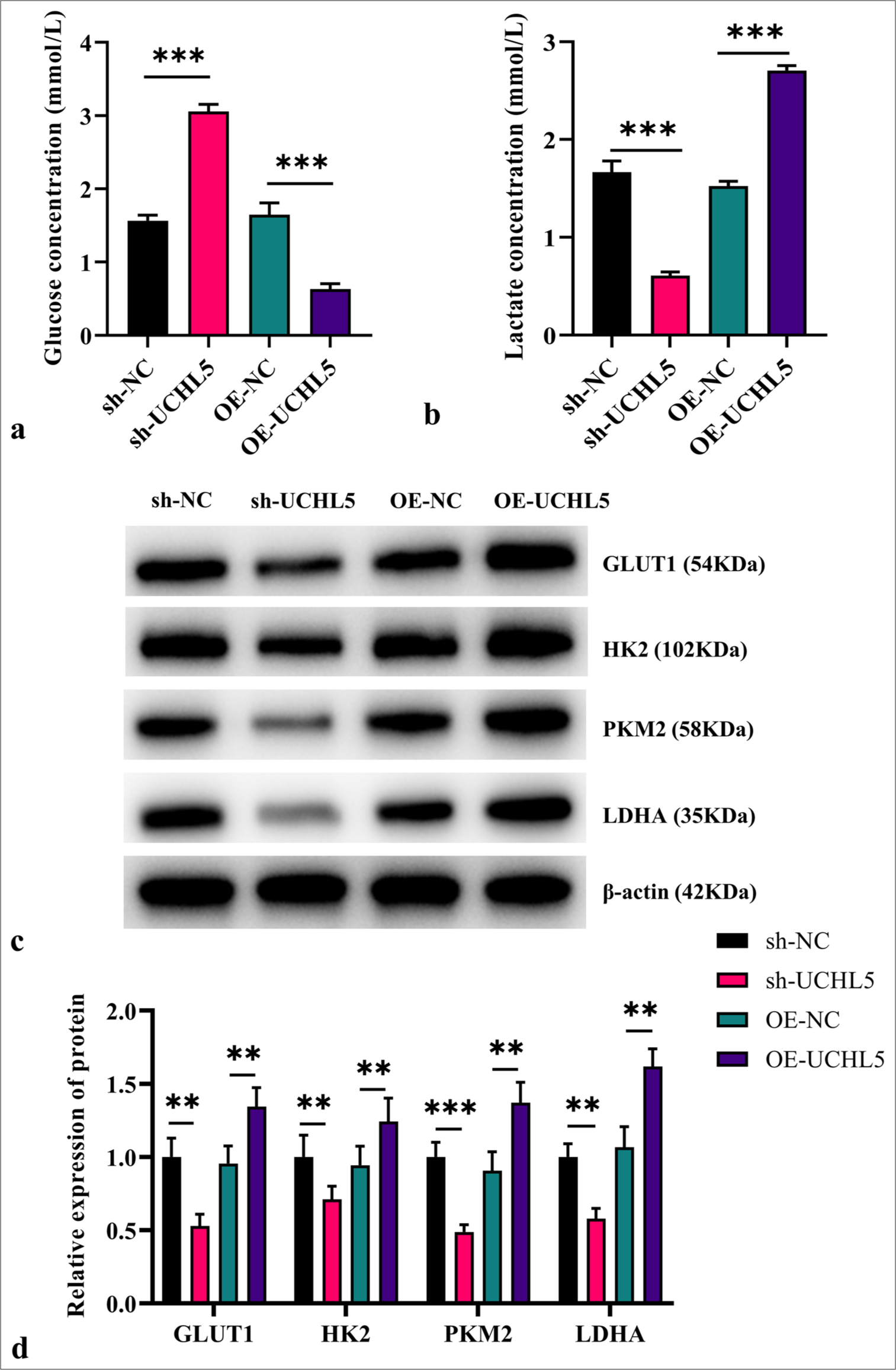
Export to PPT
UCHL5 regulated the PI3K/AKT/mTOR pathwayProteins related to the PI3K/AKT/mTOR pathway were detected after overexpression and knockdown of UCHL5 to elucidate the mechanism by which UCHL5 regulates glycolysis and malignant behavior of RCC cells. When UCHL5 was knocked down, the ratios of the phosphorylated proteins significantly decreased (P < 0.01). Treatment with the PI3K activator 740Y-P restored the reduced levels of these proteins in the sh-UCHL5 group (P < 0.01 [Figure 4a and b]). After overexpression of UCHL5, the ratios of the phosphorylated proteins markedly increased (P < 0.05). When treated with the PI3K inhibitor LY294002, the elevated levels of these phosphorylated proteins were effectively reversed (P < 0.01 [Figure 4c and d]).
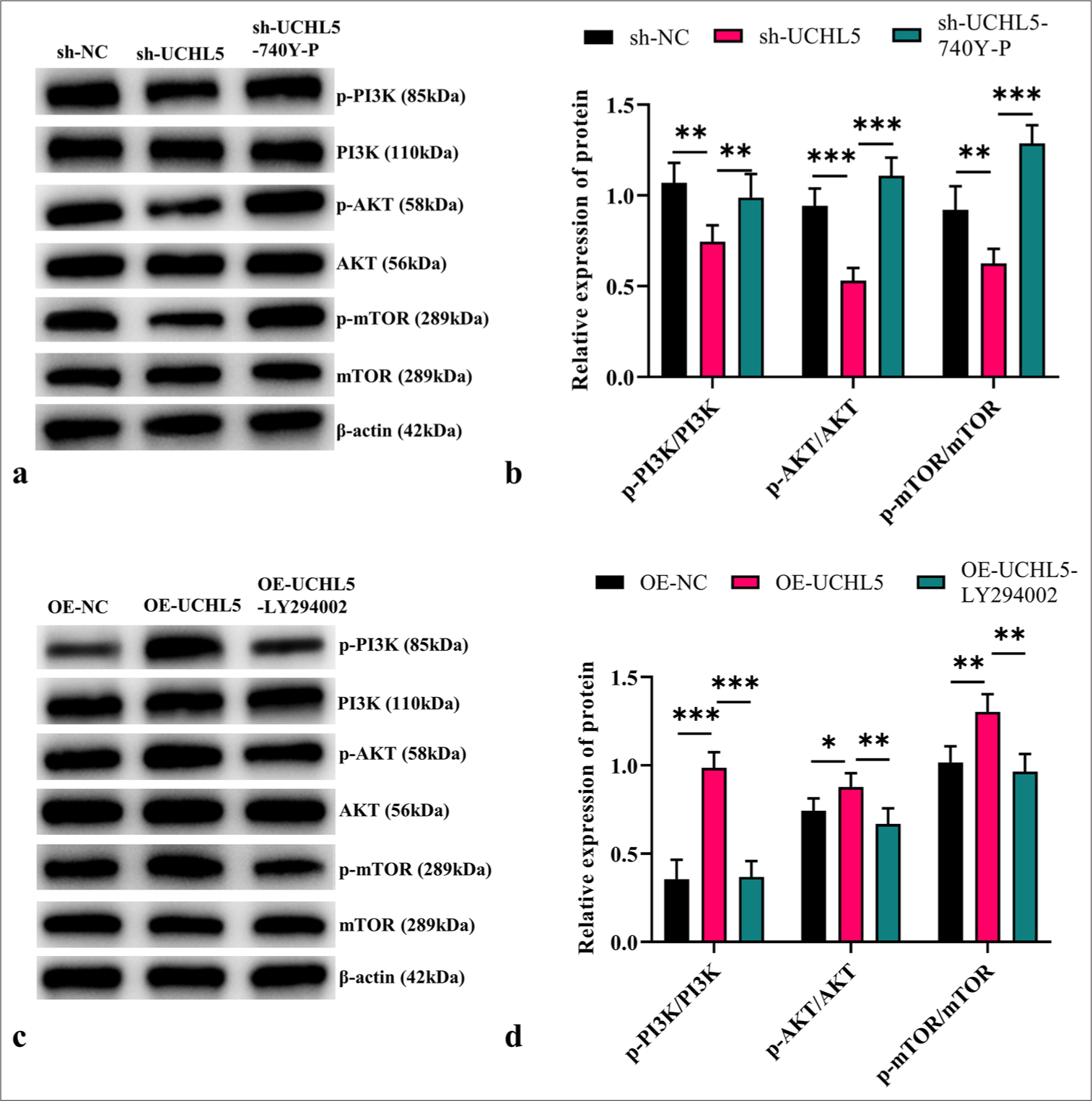
Export to PPT
UCHL5 regulated glycolysis through the PI3K/AKT/ mTOR pathwayAfter knockdown of UCHL5, glucose concentration increased (P < 0.001), whereas lactic acid production decreased (P < 0.001). However, this result was reversed in the sh-UCHL5-740Y-P group (P < 0.01). After UCHL5 was overexpressed, glucose concentration decreased, and lactic acid level significantly increased (P < 0.001). This result was reversed in the OE-UCHL5-LY294002 group (P < 0.001 [Figure 5a-d]). We further detected the expression of glycolysis-related proteins by WB. The glycolysis-related proteins were upregulated after overexpression of UCHL5 (P < 0.05). LY294002 reversed the observed upregulations of glycolysis-related proteins after UCHL5 overexpression (P < 0.01). By contrast, the depletion of UCHL5 resulted in a downregulation of glycolysis-related proteins (P < 0.01). The reduction in these protein levels caused by UCHL5 knockdown was reversed upon treatment with 740Y-P (P < 0.01) [Figure 5e-h].
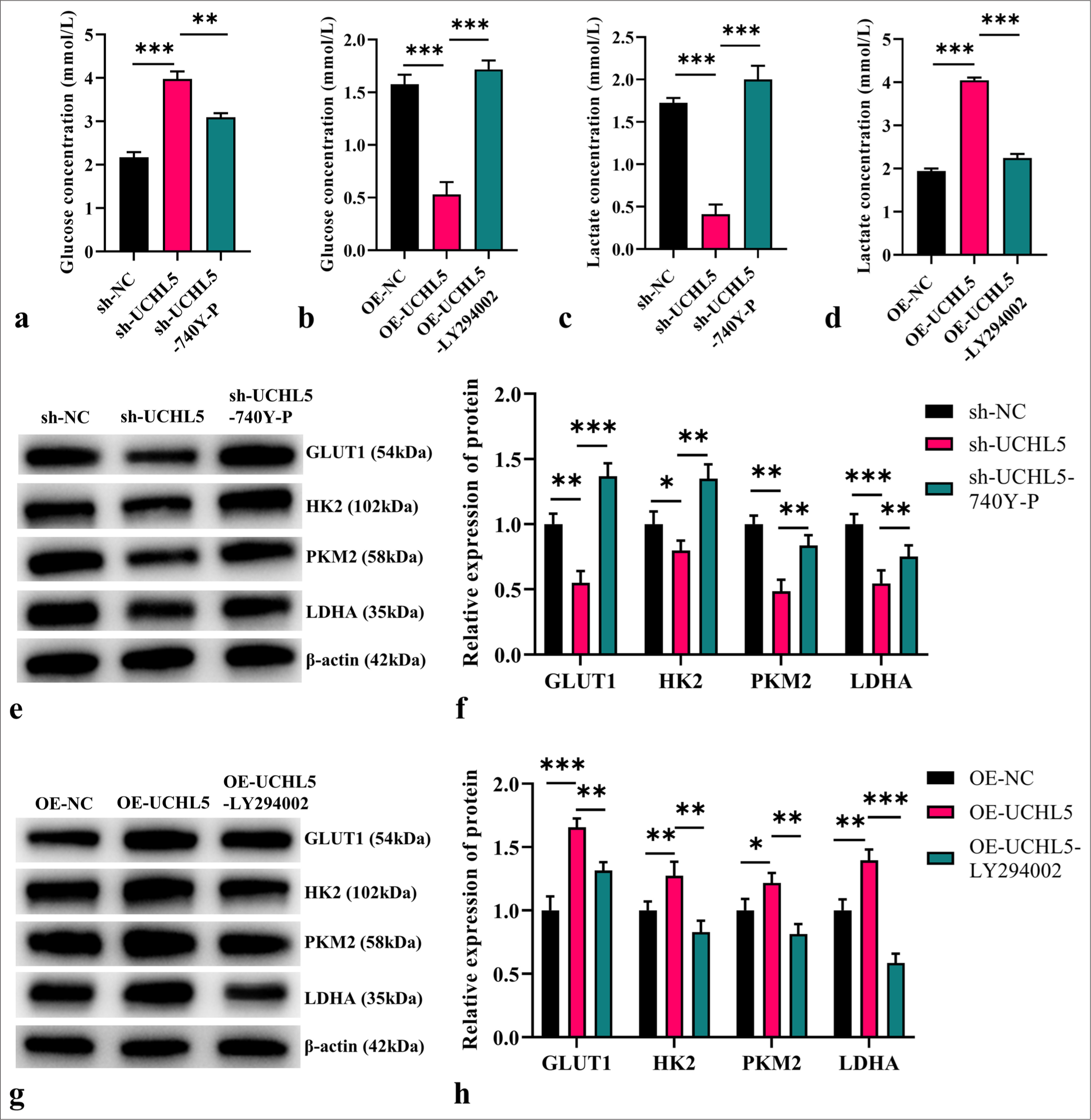
Export to PPT
UCHL5 promotes the malignancy of RCC cells through PI3K/AKT/mTOR pathwayNext, we further tested the proliferation and invasion ability of RCC cells after interference with the PI3K/AKT/ mTOR signaling pathway. The EdU assay showed that the addition of 740Y-P increased the proliferation activity of cells in the sh-UCHL5 group, and the addition of LY294002 significantly inhibited the proliferation activity of cells caused by overexpression of UCHL5 (P < 0.01 [Figure 6a-d]). The Transwell experiment showed that the addition of PI3K activator significantly increased the invasion ability of sh-UCHL5 group cells and the addition of PI3K inhibitor inhibited the increase of cell invasion caused by overexpression of UCHL5 (P<0.05 [Figure 6e-h]).
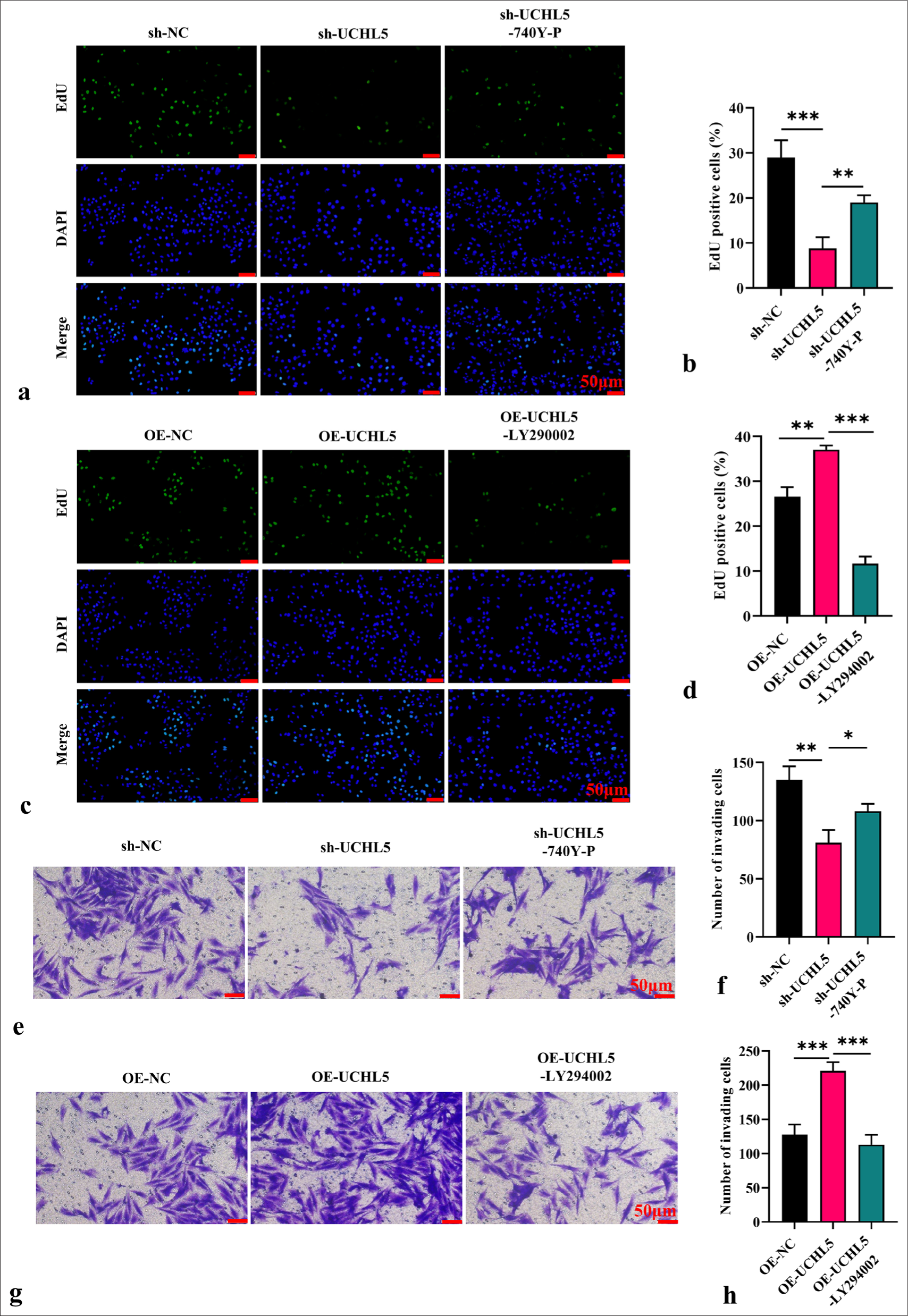
Export to PPT
Knockdown of UCHL5 reduced the glycolysis level of RCC cells and suppressed tumor development through the PI3K/AKT/mTOR pathwayLentiviral transfection was used to construct the knockdown UCHL5 cell line and its control cell line to verify the operation of UCHL5 in vivo. An animal model of transplanted tumor was established to verify the effect of knockdown UCHL5 on RCC cells in vivo. The tumor volume and weight of sh-UCHL5 group were significantly reduced (P < 0.01) [Figure 7a-c]. The IHC staining exhibited that the levels of UCHL5 and glycolytic-related proteins PKM2, LDHA, GLUT1, and HK2 were significantly decreased in sh-UCHL5 group (P < 0.01 [Figure 7d]). The IHC staining results also showed that the levels of p-PI3K/PI3K, p-AKT/AKT, and p-mTOR/mTOR in the sh-UCHL5 group were significantly decreased [Figure 8a-c]. These results provide evidence that decreasing UCHL5 expression may be a potential clinical treatment strategy for RCC.
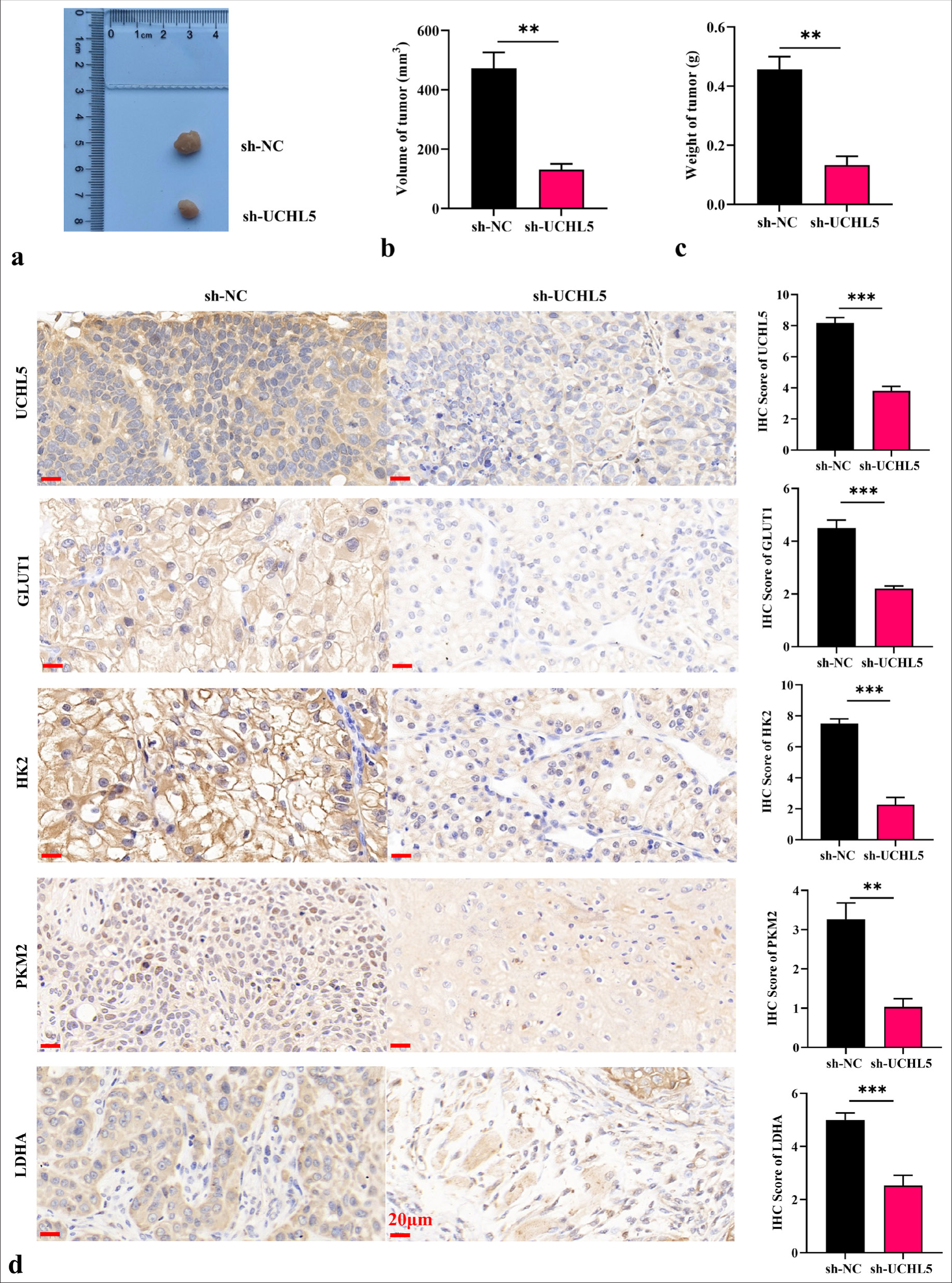
Export to PPT
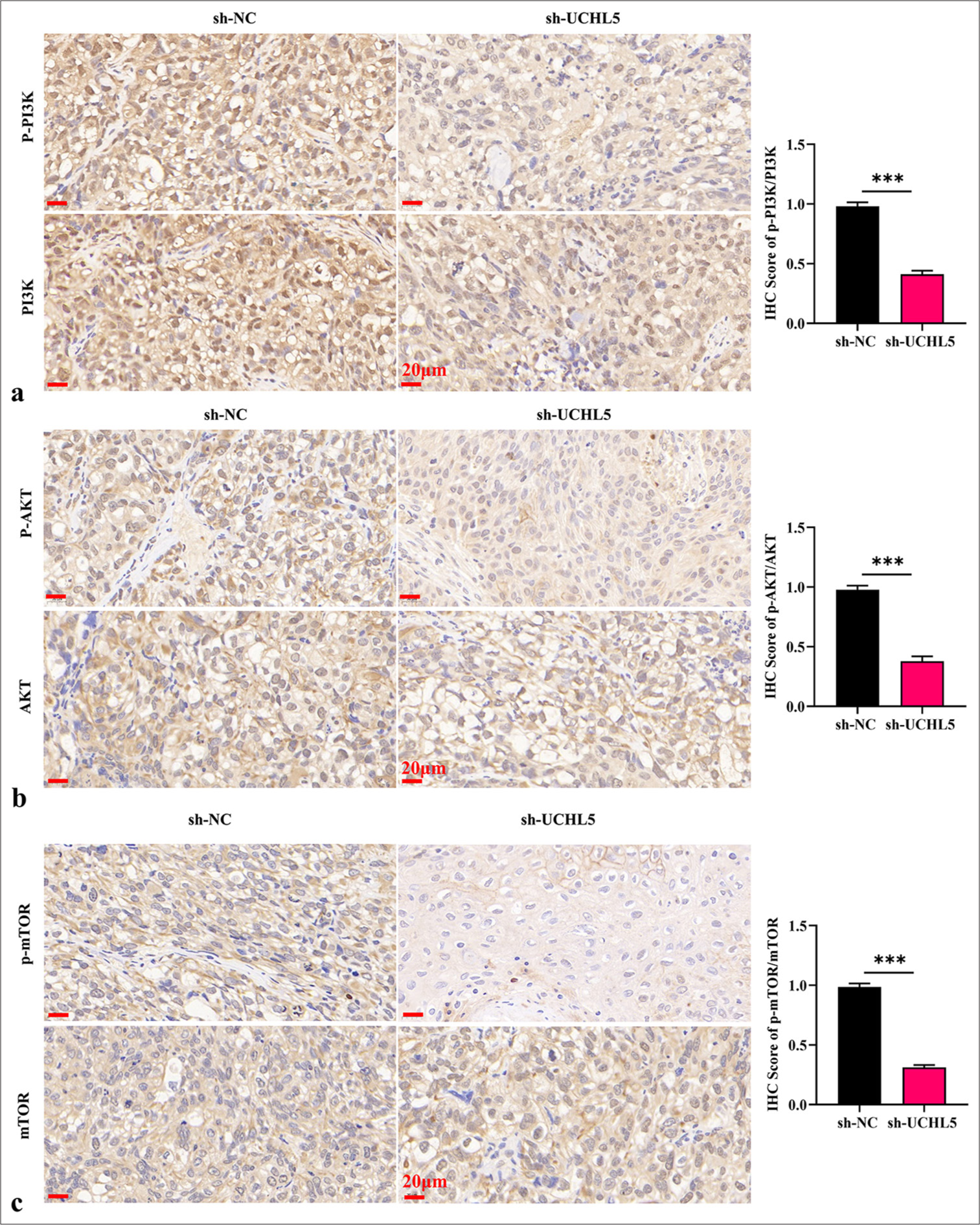
Export to PPT
DISCUSSIONRCC is a cancer occurring in the urinary system. Despite advances in treatment methods, clinical treatment is limited because of the high heterogeneity of RCC cells, their insensitivity to radiotherapy and chemotherapy, and their susceptibility to drug resistance. Patients also have a poor prognosis and a high fatality rate due to metastatic RCC.[23] Thus, investigating the mechanisms underlying RCC and identifying novel therapeutic targets have clinical importance.
Extensive research has demonstrated that UCHL5 is vital for tumor development. UCHL5 is a carboxyl-terminal hydrolase that can deubiquitinate proteins and reduce protein degradation.[20] UCHL5 can regulate RCC through the glycolysis pathway mediated by the PI3K/AKT/mTOR pathway. First, our investigation explored the expression pattern of UCHL5 in RCC cells and revealed its upregulation, consistent with the report of Zhang et al. [14] We then overexpressed and knocked down UCHL5. Overexpression of UCHL5 increased glycolysis levels and enhanced malignant behavior, suggesting that UCHL5 promotes the progression of RCC. This finding confirms the critical role of UCHL5 in tumor progression. In hepatocellular carcinoma, UCHL5 provides a proliferative advantage to tumor cells by regulating glycolytic metabolism.[20] Inhibiting the expression of UCHL5 weakens the malignant progression of ovarian cancer cells.[24]
The PI3K/AKT/mTOR pathway is crucial in the pathogenesis of RCC. PI3K activates AKT through phosphorylation, and the activated AKT increases glucose uptake and enhances the Warburg effect by activating glycolysis-related proteins.[25] mTOR is a key metabolic target for the treatment of RCC, and several studies have demonstrated its function in regulating glycolytic activity.[26,27] Blocking PI3K/AKT/ mTOR can inhibit the growth of RCC cells.[11] To explore whether UCHL5 promotes RCC through this pathway, we detected the expression of proteins related to this signaling pathway after overexpression and knockdown of UCHL5. The results showed that UCHL5 is positively correlated with the activation of this signaling pathway. Subsequently, PI3K inhibitor LY294002 and activator 740Y-P were selected for recovery experiments. Adding LY294002 to the UCHL5 overexpression group reduced the glycolysis level and cell malignant behavior. In the UCHL5 knockdown group, the addition of 740Y-P increased the glycolysis level and cell malignant behavior. Overall, inhibiting this signaling pathway reversed UCHL5 upregulation, and activating it restored the influence of UCHL5 downregulation. For instance, inhibiting the mTOR pathway can suppress the proliferation and migration of tumor cells, halt the cell cycle and promote apoptosis.[28] In bladder cancer, UCHL5 promotes tumor progression by activating the AKT/mTOR pathway and regulating downstream target proteins.[21] Ubiquitination and deubiquitination are of great significance in tumors. Interfering with UCHL5 binding to nuclear factor kappa-B (NF-κb) promotes the ubiquitination and degradation of NF-κb in gastric cancer (GC) and inhibits the proliferation and metastasis of GC cells.[29] Notably, UCHL5, as a deubiquitinating enzyme, may affect the stability and activity of key proteins in the pathway by regulating their ubiquitination levels. This mechanism provides a novel insight for understanding the relationship between UCHL5 and PI3K/AKT/mTOR pathway.
The animal experiments verified that downregulating UCHL5 reduced the glycolysis level of RCC cells and inhibited tumor growth through PI3K/AKT/mTOR. In bladder cancer, the depletion of UCHL5 suppresses tumorigenesis using a mouse in vivo model.[21] These studies provide in vivo experimental evidence for the use of UCHL5 as a therapeutic target. Given that UCHL5 activates the PI3K/AKT/mTOR pathway in RCC, the combination of UCHL5 inhibitors and mTOR inhibitors may be an effective strategy. mTOR inhibitors, such as everolimus, have been used to treat advanced RCC.[30] Our research indicates that silencing UCHL5 can reduce the activation level of the mTOR pathway. The design of small-molecule inhibitors specifically targeting UCHL5 holds great promise.[31] By simultaneously targeting UCHL5 and mTOR, we may comprehensively block the oncogenic signaling network in RCC cells. This combined treatment could enhance the anti-tumor effect and overcome resistance mechanisms associated with single-agent mTOR inhibitor therapy.
UCHL5 promotes the development of RCC through the PI3K/AKT/mTOR-glycolytic axis, but the upstream regulatory mechanism has not been fully elucidated. The effect of UCHL5 on immune cells in the tumor microenvironment and its interaction with other metabolic pathways should be further explored using in vivo models. In terms of mechanism verification, this study mainly relied on a single PI3K inhibitor and experimental methods. Potential off-target effects have not been fully ruled out. Furthermore, this study was mainly based on experiments using cell lines and animal models and lacked direct verification with clinical sample data. Future research should focus on these directions to provide a comprehensive theoretical basis and strategic support for the precise treatment of RCC.
SUMMARYUCHL5 can enhance the glycolytic pathway and facilitate RCC progression by stimulating the PI3K/AKT/mTOR pathway. Therefore, we propose that UCHL5 could serve as a novel and promising diagnostic marker and treatment target for RCC, warranting additional investigation.
Comments (0)