
Remember me
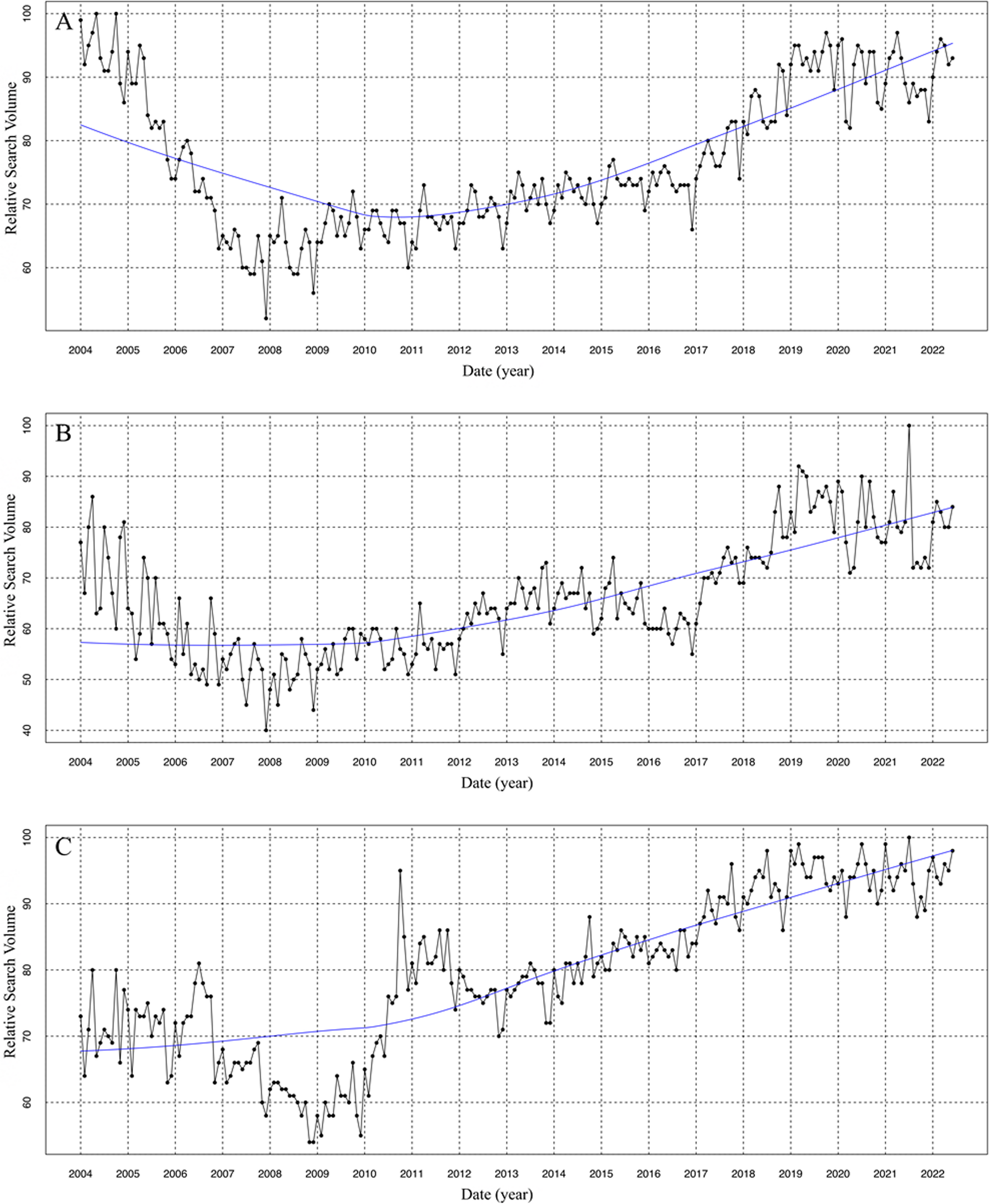
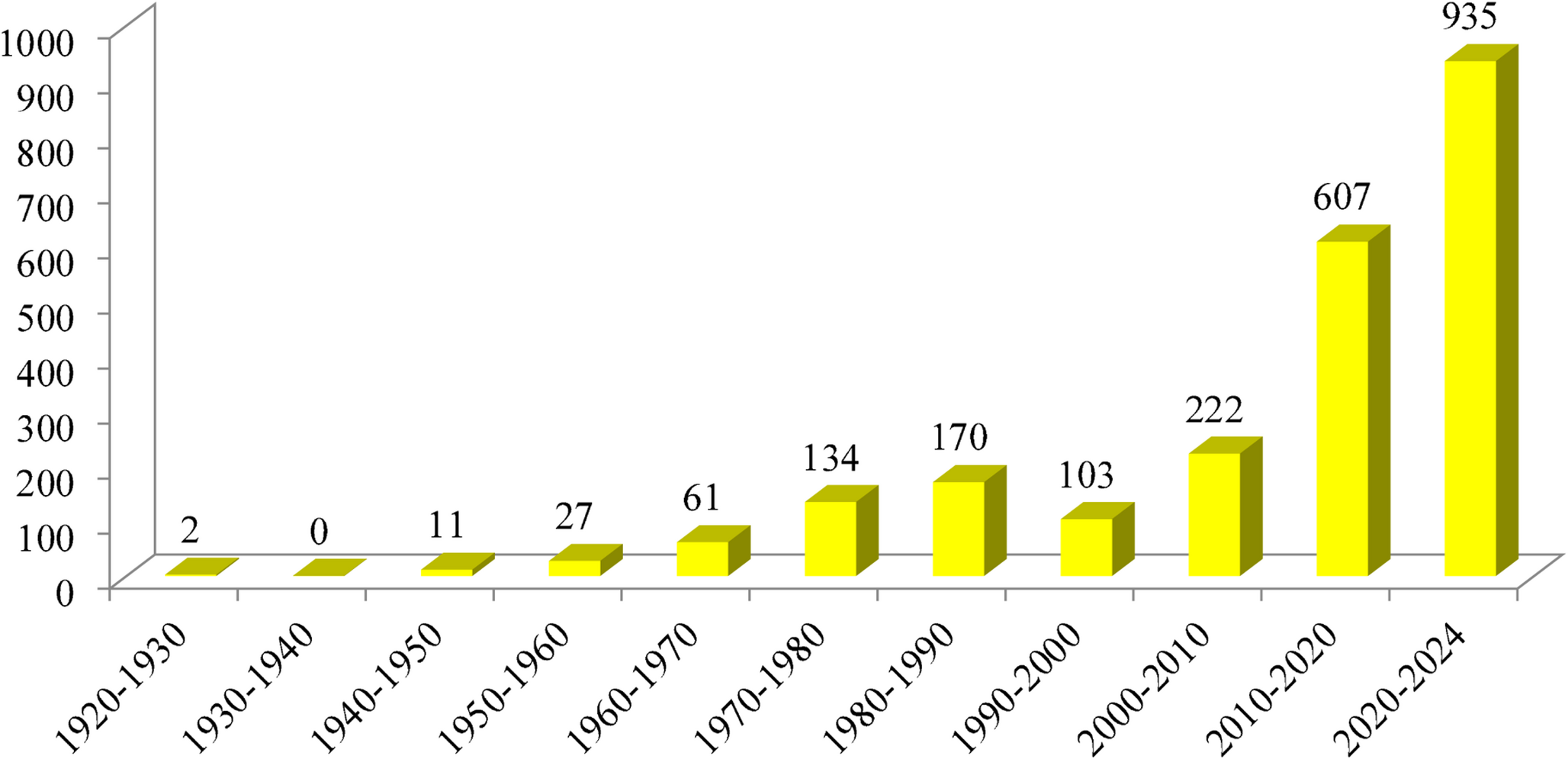
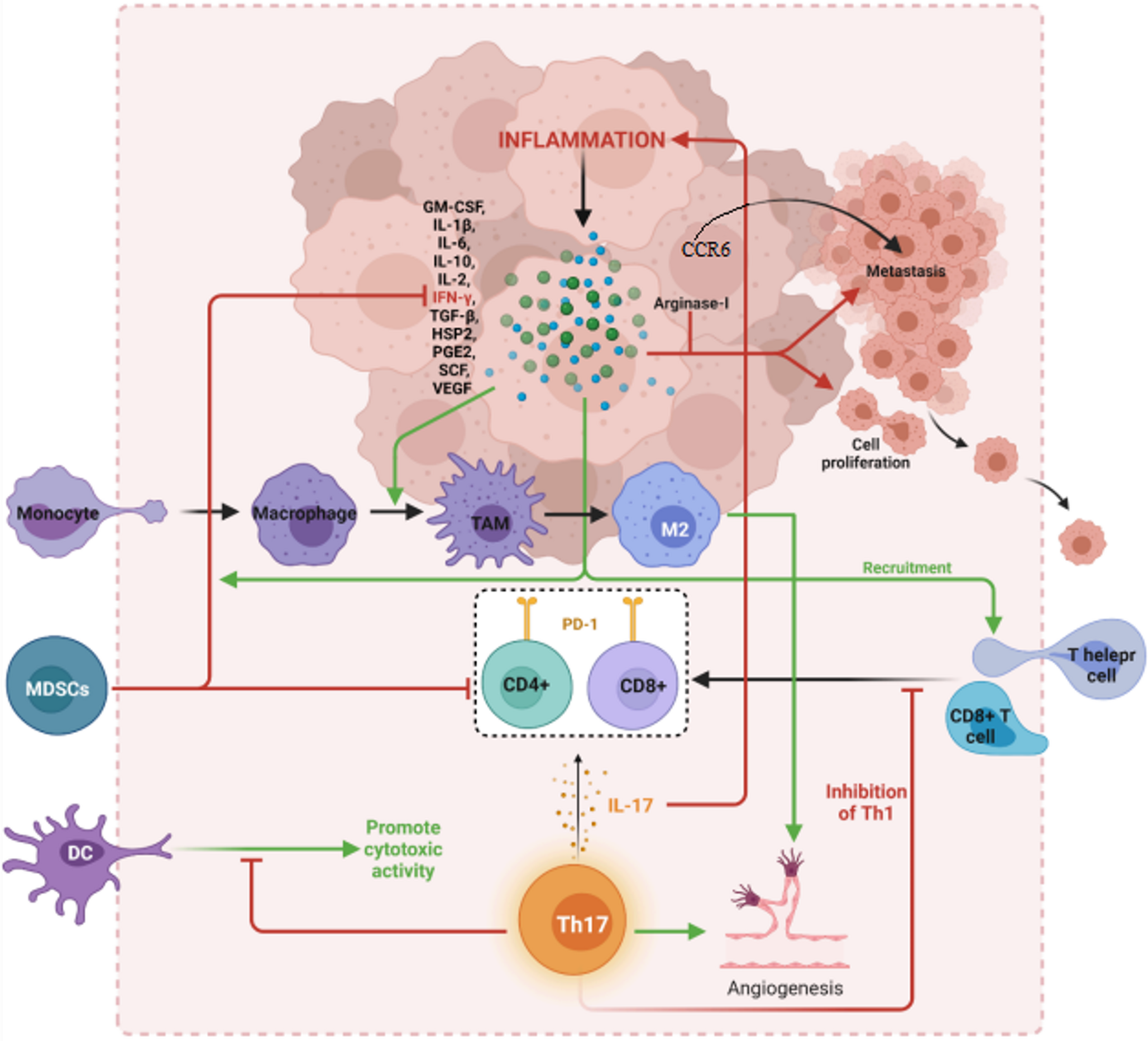
To investigate the effect of STAT6 inhibition on Treg differentiation, we induced iTregs from naive CD4⁺ T cells derived from WTGFP and STAT6−/−GFP mice. The process for T-cell isolation, activation, iTreg induction, and expansion is outlined in Fig. 1A. After a 5-day culture period, the cells were stained with anti-CD4 and anti-CD25 antibodies and analyzed by FCM to evaluate the expression of Treg-specific surface markers. As anticipated, over 85% of the cells successfully differentiated into CD4⁺Foxp3⁺CD25⁺ iTregs (Fig. S1).
Fig. 1
STAT6 inhibition promotes the stabilization of iTreg differentiation in vitro. Naive CD4+ T cells were isolated from the spleens of 8–10-week-old WT or STAT6−/− Foxp3GFP mice and differentiated into inducible Tregs (iTregs) over five days (A). During this time, the cells were activated with anti-CD3 and anti-CD28 antibodies, TGF-β and IL-2. After five days, TGF-β and the anti-CD3/CD28 antibodies were removed from the culture, and the iTregs were expanded. On day 7, the iTregs were treated with AS1517499 (AS) or left untreated for 72 h. Following this treatment, the cells were harvested and analyzed using flow cytometry (FCM). Representative FCM plots and percentages of CD4+Foxp3+ cells are shown in (B), CD4+Foxp3+CD25+ cells in (C), CD4+Foxp3+PD-1+ cells in (D), and CD4+Foxp3+CTLA-4+ cells in (E). The mean fluorescence intensity (MFI) of Foxp3 (F) and CD25 expression (G) in WT or STAT6−/− iTregs and those treated with the STAT6 inhibitor (AS-iTregs) are also displayed. n = 3 per group; the statistical significance was determined using One-way ANOVA and Bonferroni’s multiple comparison test; *p < 0.05, **p < 0.01, and ***p < 0.001; 4 experimental replicates)
To determine whether pharmacological inhibition of STAT6 signaling with AS1517499 promotes iTreg stability in a manner similar to that observed in STAT6−/− mice, we cultured iTregs in the presence of IL-2 and 100 nM of AS1517499 for 72 h (Fig. 1A). To confirm the direct effect of AS1517499 on STAT6 activity in iTregs, we evaluated both total STAT6 and phosphorylated STAT6 (pSTAT6) levels following treatment. Our results demonstrated that 100 nM AS1517499 effectively reduced STAT6 phosphorylation without altering total STAT6 expression (Fig. S2), indicating sufficient inhibition of STAT6 signaling under these conditions. Following ten days of expansion, the frequency of the CD4⁺Foxp3⁺CD25⁺ cells significantly declined to below 59% ± 4 in the WT-iTregs. In contrast, the frequency of iTregs generated with STAT6 pharmacological inhibition (AS-iTregs) remained stable at approximately 80% ± 1.4 after the same period (Fig. 1B, C), with negligible loss in Foxp3 and CD25 expression. As expected, in STAT6−/− animals, the frequency of CD4⁺Foxp3⁺CD25⁺ cells remained consistently high, reaching up to 75% ± 2, and this effect was independent of STAT6 inhibitor treatment (Fig. 1B, C).
The expression of activation and proliferation markers was also accompanied by a marked increase in the co-expression of PD-1 (35.2 ± 2% vs. 30.6 ± 0.5%, p < 0.05) and CTLA-4 (33.7 ± 2% vs. 21% ± 2, p < 0.05) in WT AS-iTregs compared to WT-iTregs (Fig. 1D, E). Furthermore, WT AS-iTregs showed the highest proportions of Foxp3⁺ and CD25⁺ cells, supporting enhanced Treg stability during in vitro expansion (Fig. 1F, G). Together, these findings indicate that pharmacological inhibition of STAT6 using AS1517499 significantly promotes iTreg differentiation and stability in vitro, recapitulating the phenotype observed in STAT6-deficient mice.
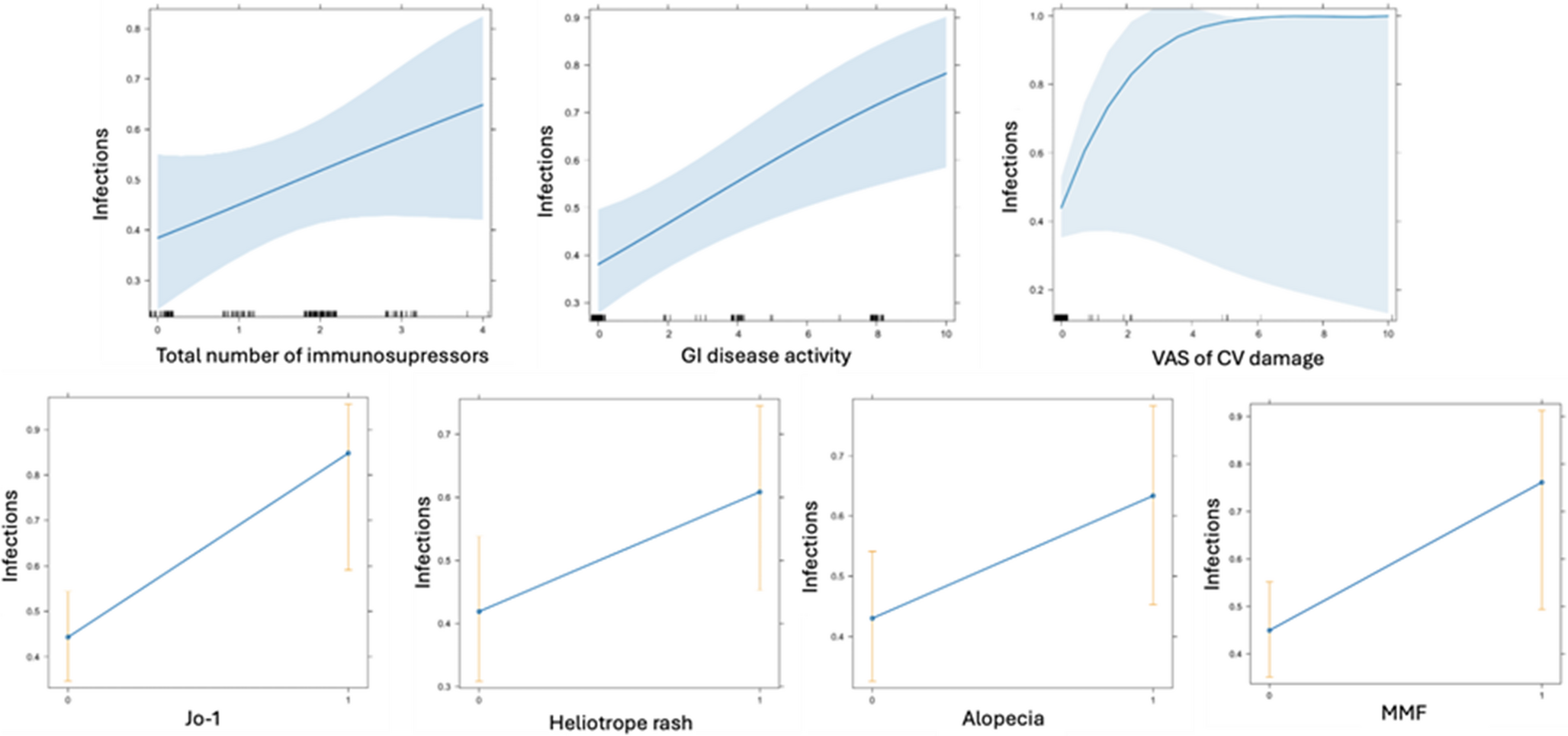
iTregs induced under STAT6 inhibition remain stable under inflammatory conditions and exhibit enhanced suppression of CD4+ T cells in vitroTo assess whether iTregs generated in the absence or inhibition of STAT6 remain stable under inflammatory conditions, we cultured iTregs with the pro-inflammatory cytokine IL-6 for 3 days during the expansion phase (Fig. 2A). In WT iTregs, the frequency of CD4⁺Foxp3⁺ cells significantly decreased in the presence of IL-6 compared to WT iTregs generated under pharmacological STAT6 inhibition (WT AS-iTregs) (55.9 ± 6% vs. 78.0 ± 0.5%, p < 0.001) (Fig. 2 B,C). In contrast, STAT6−/− iTregs showed no significant loss of CD4⁺Foxp3⁺ cells upon IL-6 exposure, with comparable frequencies observed between STAT6−/− iTregs and STAT6−/− AS-iTregs (70.3 ± 4.2% vs. 77.5 ± 1.4.%, NS), indicating that STAT6 signaling deficiency supports phenotypic stability of iTregs even under cytokine-induced stress (Fig. 2B, C).
Fig. 2
Blocking STAT6 preserves iTreg identity and potentiates suppression despite inflammatory signals. A Naive CD4+ T cells were isolated from the spleens of 8–10-week-old WT or STAT6−/− Foxp3GFP mice and differentiated into inducible Tregs (iTregs) over five days. During this time, the cells were activated with anti-CD3 and anti-CD28 antibodies, TGF-β and IL-2. After five days, TGF-β and the anti-CD3/CD28 antibodies were removed from the culture, and the iTregs were expanded in the presence of IL-2, AS1517499 (AS), IL-6, or all-trans retinoic acid (atRA) for 72 h. Following this treatment, the cells were harvested and analyzed using flow cytometry (FCM). B Representative FCM plots and C percentages of CD4+Foxp3+ cells. Statistical significance was determined using One-way ANOVA and Bonferroni’s multiple comparison test; *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001; a total of 3 experimental replicates were performed. D Histograms illustrate the proliferation of CD4+ responder T cells (Tresp) in the absence or presence of iTregs. iTregs were in vitro generated under Treg polarization conditions from WT mice or treated with 100 nM AS1517499 for 72 h during expansion (AS-iTregs) and FACS cell-sorted CD4+Foxp3+ cells. Tresp were treated as described in the materials and methods. Numbers indicate the percentage of proliferating cells when iTregs and Tresp cells are co-cultured at different ratios. Graph showing the percentage of total divided CD4+ cells at varying iTreg: Tresp ratios. The lymphocyte population was gated based on forward scatter (FSC) and side scatter (SSC) characteristics and further subgated for CD4 expression. Five thousand events were captured from each subgate. The graph shows the mean ± SEM from two experiments with similar results. Statistical significance was determined using a two-tailed unpaired Student’s t-test (*p < 0.05, **p < 0.01, 3 experimental replicates)
Previous studies have shown that all-trans retinoic acid (atRA), a vitamin A derivative, significantly enhances the phenotypic and functional development of TGF-β–induced iTregs and supports their maintenance [11]. To determine whether combining STAT6 inhibition with atRA further improves iTreg stability and function, we co-administered atRA and the STAT6 inhibitor AS1517499 during iTreg expansion (Fig. 2B). However, no significant increase in the frequency of CD4⁺Foxp3⁺ cells was observed in either WT or STAT6−/− cells treated with the combination, suggesting that atRA and AS1517499 do not exert a synergistic or additive effect.
We observed that pharmacological STAT6 inhibition increased the expression of Foxp3, CD25, PD-1, and CTLA-4 in WT AS-iTregs compared to WT iTregs at 10 days of expansion. Therefore, we decided to evaluate whether iTregs developed under these conditions exhibit a superior suppressive capacity.
To assess the suppressive ability of CD4+Foxp3+ cells, we sorted these cells from the cultures and tested them against naïve splenocytes that were stimulated with α-CD3. The naïve splenocytes were exposed to different ratios of iTreg cells. Upon stimulation with α-CD3, the total splenocytes exhibited strong proliferation of CD4+ T cells, as expected (Fig. 2D). Notably, AS-iTregs demonstrated a significantly enhanced suppressive capacity compared to CTR-iTregs, particularly at higher dilutions (1:3 and 1:7) (Fig. 2D). Collectively, these findings suggest that blocking STAT6 signaling not only stabilizes the iTreg phenotype but also enhances their immunosuppressive function.
STAT6 inhibition results in higher levels of Foxp3, CTLA-4, PD-1, IL-10, and TGF-β mRNA expression during iTreg expansionOur findings indicated that iTregs generated from WT mice treated with 100 nM AS1517499 (AS-iTregs) exhibited higher levels of Foxp3 and CD25 during expansion on day 10 of culture compared to control iTregs. Therefore, we further evaluated the mRNA expression of genes considered hallmarks of Tregs. AS-iTregs showed increased relative mRNA expression of Foxp3 and CTLA-4 than CTR iTregs. Both Foxp3 and CTLA-4 are crucial for the function of Tregs in vivo (Fig. 3A, B). A significant elevation in PD-1 mRNA was also observed in AS-iTregs in contrast to control iTregs (Fig. 3C). Notably, AS-iTregs demonstrated a substantial increase, reaching 7–9 times higher levels of CTLA-4 and PD-1 markers compared to iTregs.
Fig. 3
STAT6 modulates the expression of key mRNA programs during iTregs expansion. WT iTregs were expanded as in Fig. 1A and cultured with vehicle or 100 nM AS1517499 (AS-iTregs). At the indicated time point, total RNA was extracted and qRT-PCR performed. The resulting graphs illustrate relative mRNA expression levels for various markers: Foxp3 (A), CTLA-4 (B), PD-1 (C), IL-10 (D), TGF-β (E), IL-6 (F), IL-1β (G), and DNMT1 (H). Data are expressed as mean ± SEM from two independent experiments that yielded comparable results, assessed using a two-tailed unpaired Student’s t-test, with significance indicated as *p < 0.05, **p < 0.01, and ***p < 0.001, 3 experimental replicates
We assessed the mRNA expression levels of the cytokines IL-10 and TGF-β, finding that AS-iTregs exhibited a notable increase in the mRNA levels of IL-10 (Fig. 3D) and TGF-β (Fig. 3E) compared to regular iTregs. It is important to note that IL-6 and IL-1β are cytokines that help regulate the balance between Tregs and Th17 cells. IL-6 inhibits TGF-β-induced Tregs differentiation and can downregulate Foxp3, while IL-1β plays a critical role in the early differentiation of Th17 cells [4, 12]. Interestingly, the iTregs expanded with STAT6 inhibitor showed more than a 50% reduction in IL-6 and IL-1β expression levels compared to iTregs (Fig. 3F, G).
TGF-β promotes Foxp3 expression by inhibiting the activity of DNA methyltransferase 1 (DNMT1), which is responsible for maintaining DNA methylation patterns, including those in the regulatory regions of the Foxp3 gene [13]. We also observed that STAT6 inhibition led to a significant reduction in DNMT1 expression in AS-iTregs compared to control iTregs (Fig. 3H). Removing TGF-β from the culture typically results in iTreg instability and increased DNMT1 expression. However, the reduced DNMT1 expression observed in AS-iTregs after 10 days of culture, even in the absence of TGF-β, may contribute to the enhanced stability of Foxp3 expression.
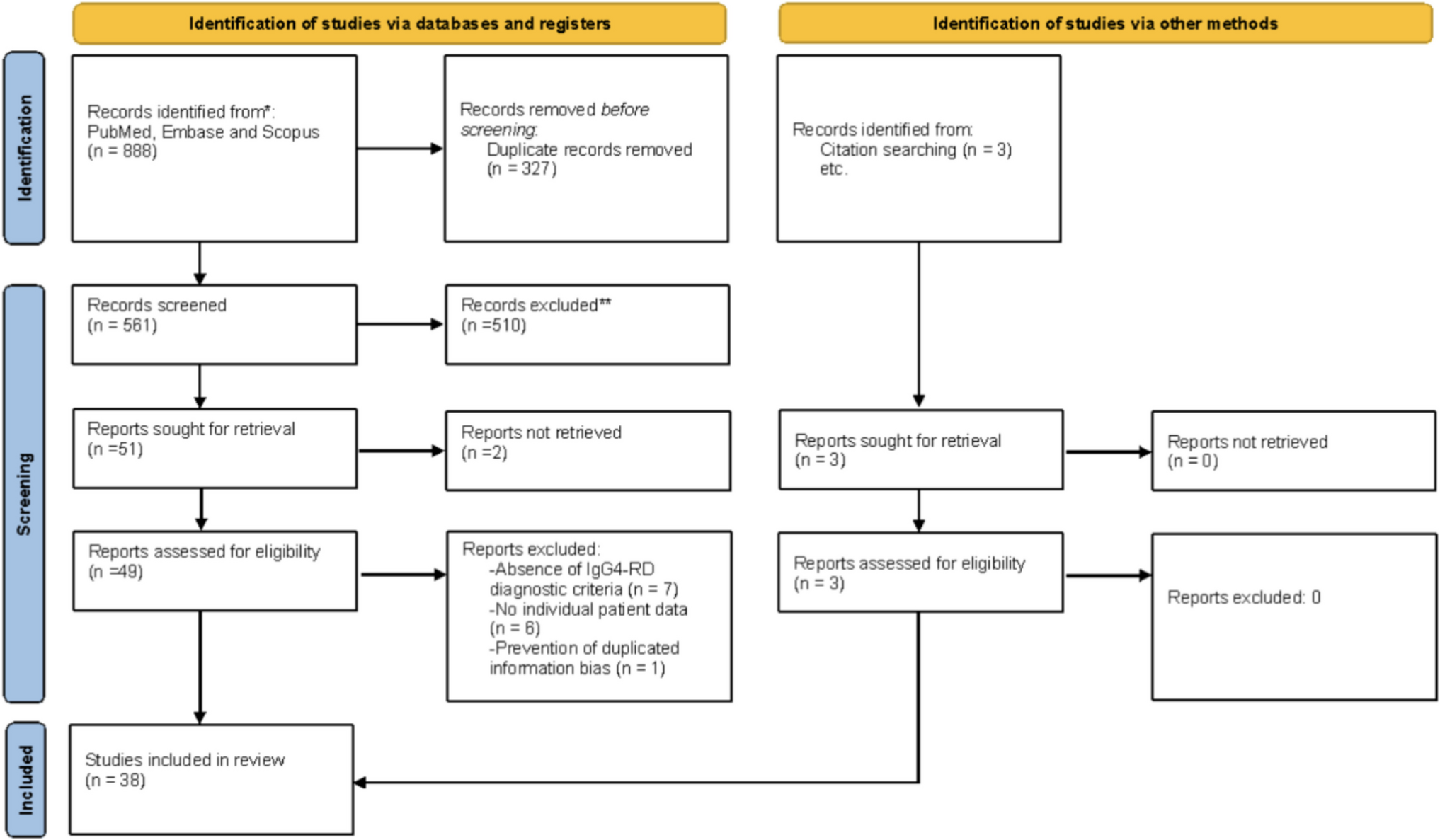
STAT6 inhibition is crucial for maintaining potent Treg function to suppress colitisTo assess whether iTregs generated in the absence or inhibition of STAT6 remain stable under in vivo inflammatory conditions, we intravenously transferred FACS-sorted control iTregs, AS-iTregs, or STAT6−/− iTregs (3 × 105 cells) into mice during AOM/DSS-induced colitis (Fig. 4A). Following DSS treatment, mice receiving either CTR iTregs, AS-iTregs, or STAT6−/− iTregs exhibited less weight loss (Fig. 4B) and demonstrated reductions in the overall disease activity index (DAI) score (Fig. 4C).
Fig. 4
Attenuation of colonic inflammation by adoptive transfer of AS1517499 (AS)-iTregs. A Induced regulatory T cells (iTregs) were sorted and intravenously (i.v.) injected into mice subjected to azoxymethane (AOM)/dextran sodium sulfate (DSS)-induced colitis. Colitis was initiated with a single intraperitoneal injection of AOM followed by administration of 2% DSS in drinking water for 7 days, repeated for one cycle of DSS administration. The vehicle group received 100 μL of saline, while control iTregs, AS-iTregs, and STAT6−/− iTregs groups received 3 × 105 iTregs via i.v. injection on day 13. Mice that received only normal drinking water served as healthy controls. B Body weight changes, expressed as a percentage of baseline, and C disease activity index (DAI) were monitored twice weekly throughout the experiment. D On day 40, colon tissues were harvested and analyzed by hematoxylin and eosin (H&E) staining to assess histopathology and by immunohistochemistry for Foxp3, iNOS, and CD8 expression. E Histological scores, F goblet cell counts (via Alcian blue staining), and G spleen weights were quantified. Data are presented as mean ± SEM (n = 3 mice per group), representative of two independent experiments. Statistical analysis was performed using one-way ANOVA with Bonferroni’s post hoc test. *p < 0.05, **p < 0.01, and ***p < 0.001
Hematoxylin and eosin (H&E) staining revealed that vehicle-treated mice subjected to AOM/DSS exhibited severe colonic damage, characterized by destruction of the intestinal epithelial structure, crypt loss, inflammatory cell infiltration, and marked edema (Fig. 4D), which was reflected in significantly elevated pathological scores (Fig. 4E). In contrast, mice treated with AS-iTregs showed substantial improvement in histopathological features, with restoration of crypt architecture and reduced inflammation, comparable to the effects observed with STAT6−/− iTregs. Conversely, mice receiving vehicle alone or control iTregs (iTregs) continued to display pronounced crypt disruption, persistent inflammatory infiltrates, and splenomegaly (Fig. 4E–G).
To assess intestinal barrier integrity, Alcian blue staining was performed to detect acid mucins. The gut tissues of mice in the vehicle and control iTregs groups showed a significant reduction in goblet cell numbers. In comparison, AS-iTregs and STAT6−/− iTregs transfection significantly increased the number of goblet cells (Fig. 4F). These findings indicate that AS-iTregs treatment can effectively rescue AOM/DSS-induced gut damage during chronic colitis.
To assess the immunological impact of these transfection therapies, we analyzed immune cell populations implicated in tissue damage and immune regulation in IBD. Treg infiltration was assessed by immunohistochemistry for Foxp3⁺ cells. Only sparse Foxp3⁺ cells were observed in the colonic tissue of mice receiving vehicle, control iTregs, or STAT6−/− iTregs. In contrast, AS-iTreg-treated mice showed a marked increase in Foxp3⁺ Treg infiltration in the colon (Fig. 4D), indicating enhanced local Treg recruitment and potential immunoregulatory activity.
Given that CD8⁺ effector T cells play a key role in epithelial injury during IBD, we also assessed CD8⁺ cell infiltration in colonic tissues. Notably, AS-iTreg therapy was associated with a reduction in CD8⁺ T cell accumulation, whereas substantial CD8⁺ cell infiltration was observed in the colons of mice treated with vehicle, control iTregs, or STAT6−/− iTregs (Fig. 4D).
Additionally, we examined inducible nitric oxide synthase (iNOS) expression, a pro-inflammatory enzyme upregulated during colitis, particularly in inflamed epithelium and infiltrating immune cells. As expected, iNOS⁺ cells were markedly increased in the colons of vehicle-treated mice. Interestingly, no reduction in iNOS⁺ cell numbers was observed in mice treated with control iTregs or AS-iTregs. A decrease in iNOS expression was only evident in the STAT6−/− iTregs group (Fig. 4D). It is important to note, however, that while iNOS upregulation is a hallmark of inflammation in colitis, its role in disease pathogenesis is complex and may vary depending on the stage and subtype of the disease.
AS-iTregs suppress inflammatory responses without exacerbating tumor development in CACPD-1 and CTLA-4 can have context-dependent roles in immune regulation, potentially contributing to both immune suppression and tumor progression. To assess whether STAT6 inhibition-induced iTregs (AS-iTregs) could exert excessive immunosuppression or promote tumor growth, we compared the effects of adoptively transferred control iTregs and AS-iTregs in the AOM/DSS-induced colitis-associated cancer (CAC) model (Fig. 5A). iTregs were administered intravenously in three adoptive transfers during the inflammatory phases of CAC development. Mice were sacrificed on day 71, a time point at which adenocarcinomas were well established. This experimental design allowed us to assess the potential impact of AS-iTregs on tumor progression under chronic inflammatory conditions.
Fig. 5
AS-iTregs alleviate inflammation without affecting tumor progression in a CAC model. A iTregs were sorted and injected intravenously (i.v.) into mice subjected to AOM/DSS-induced CAC. The vehicle group received 100 μL of saline, while the control iTregs and AS-iTregs groups were injected i.v. with 3 × 105 FACS-sorted cells on days 13, 27, and 41. Mice given normal distilled water served as controls. Effects of iTreg transfer on disease progression: B changes in body weight; C disease activity index; D, E colon length; F number of tumors; G tumor area; and H histological sections of inflamed colons at day 71. I Survival rates were recorded daily following CAC initiation. Data are presented as mean ± SEM for 6 mice per group. Similar results were observed in two independent experiments. One-way ANOVA and Bonferroni’s multiple comparison tests (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001)
AOM/DSS treatment in the vehicle group resulted in significant body weight loss and increased DAI scores. In contrast, mice treated with iTregs or AS-iTregs exhibited a protective effect, including reduced weight loss and improved diarrhea and intestinal bleeding (Fig. 5B, C).
Inflammation associated with colitis typically shortens the colon. AS-iTregs treatment significantly preserved colon length compared to vehicle-treated animals (Fig. 5D, E). As expected, all mice in the vehicle group developed colon tumors. In contrast, both iTregs and AS-iTregs significantly reduced tumor incidence, though neither group showed an effect on tumor size (Fig. 5 F, G). While the colon tissues of iTregs and AS-iTregs-treated mice displayed reduced inflammation, disrupted crypt architecture was still evident, and no differences were observed in total histological score (Fig. 5H).
Notably, a significant difference in survival was observed between mice with or without iTreg treatment (Fig. 5H). However, no differences were observed between iTregs and AS-iTregs-treated mice. Our results demonstrate that STAT6 inhibition enhances iTreg differentiation and mitigates inflammation, though this immunomodulation alone is insufficient to prevent CAC development, highlighting the complexity of tumor-immune interactions in chronic inflammation.
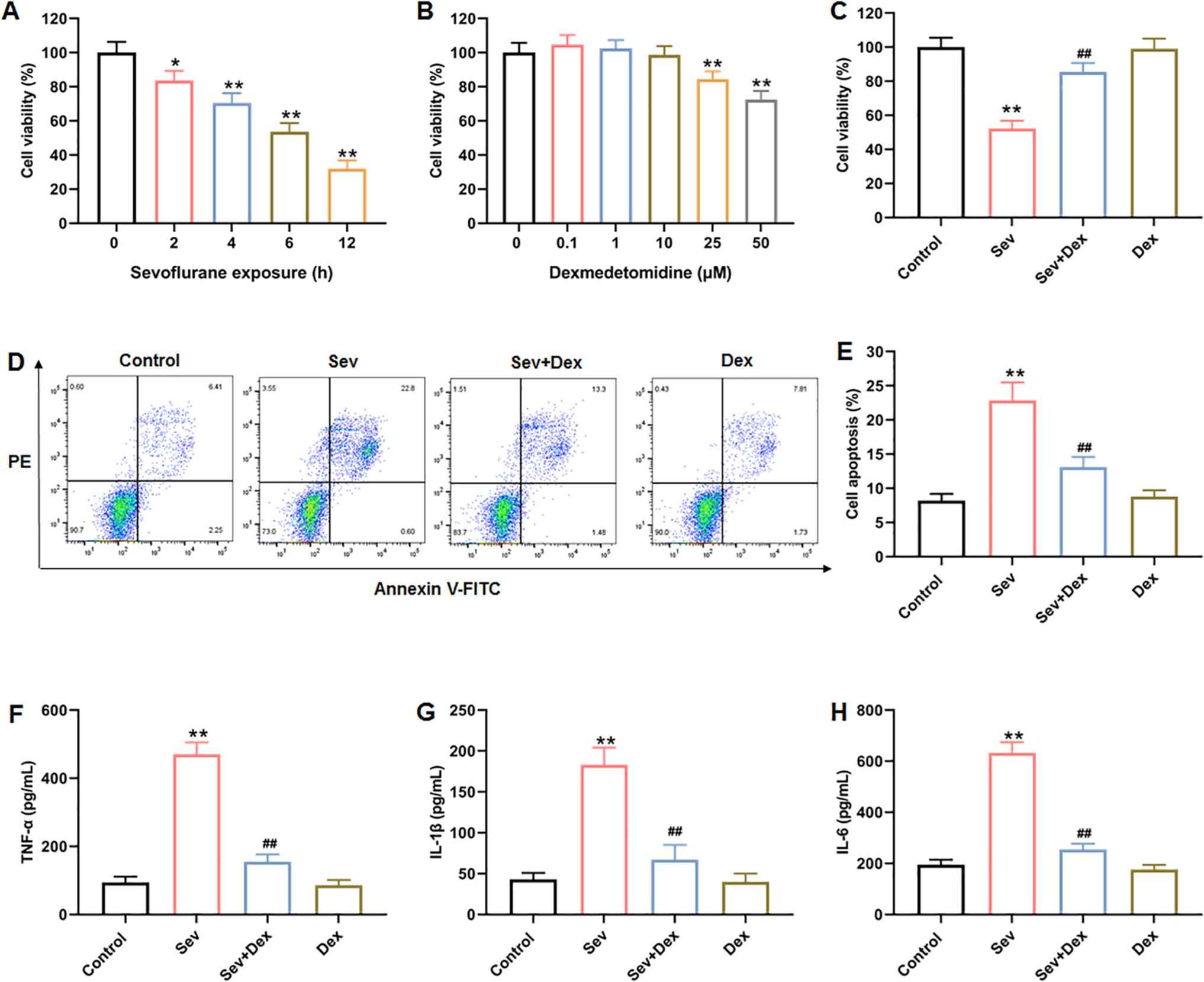
Pharmacological STAT6 inhibition promotes Treg-mediated immune regulation in acute colitisBuilding on our in vitro findings that STAT6 inhibition enhances the stability and suppressive function of iTregs, we next investigated whether in vivo administration of the STAT6 inhibitor AS1517499 could similarly enhance Treg activity and attenuate inflammation in a model of acute colitis. To this end, we utilized the DSS-induced acute colitis model in WT Foxp3EGFP mice (Fig. 6A). Consistent with previous reports, mice treated with DSS and vehicle exhibited progressive weight loss and a marked increase in DAI over time. In contrast, mice treated i.p. with AS1517499 displayed significantly reduced weight loss and DAI, confirming the anti-inflammatory effects of STAT6 inhibition (Fig. 6B and C), in line with our earlier findings [14].
Fig. 6
Pharmacological STAT6 inhibition promotes Treg-mediated immune regulation in acute colitis. A Acute colitis was induced by administering 4% DSS in drinking water for 7 days. The vehicle group received 100 μL of DMSO, while the AS1517499 (AS) group was treated intraperitoneally with 10 mg/kg of AS on days 1, 3, and 5. A separate group receiving normal drinking water served as healthy controls. B Body weight change (expressed as a percentage of baseline) and C disease activity index (DAI) were monitored daily. Representative flow cytometry plots and frequencies of D CD4⁺Foxp3⁺ cells, E CD4⁺Foxp3⁺CD25⁺ cells, F IFN-γ⁺, and G PD-1⁺ expression in CD4⁺Foxp3⁺ Tregs from mesenteric lymph nodes (MLNs). Data are shown as mean ± SEM (n = 4–5 mice per group) and are representative of two independent experiments. Statistical significance was determined using one-way ANOVA followed by Bonferroni’s post hoc test (*p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001). In panels B and C, asterisks indicate differences between the DSS + AS group and all other groups
However, in our previous study, we did not evaluate whether this reduced inflammation was associated with enhanced Treg proliferation or function. Here, we demonstrate that AS1517499 treatment during acute colitis prevents the decline in the frequency of CD4⁺Foxp3⁺ Tregs (Fig. 6D) and promotes the maintenance of activated Treg subsets, including CD4⁺Foxp3⁺CD25⁺ cells (Fig. 6E), IFN-γ–expressing Tregs (CD4⁺Foxp3⁺IFN-γ⁺; Fig. 6 F), and PD-1–expressing Tregs (CD4⁺Foxp3⁺PD-1⁺; Fig. 6G). These results provide novel evidence that pharmacological inhibition of STAT6 in vivo enhances Treg expansion and activation, while supporting their suppressive capacity under inflammatory conditions.
Comments (0)