
Remember me

K1 Causality & Beyond: Neuroscience in the Data-Deluge Era
1Dept. of Neurobiology, University of Pennsylvania
*Email: kording@upenn.edu
Neuroscience is thriving in an era of unprecedented data. Advances in recording, imaging, and machine learning have opened new frontiers—but understanding how brain activity causes behavior remains elusive. Traditional inference struggles in vast hypothesis spaces, and many neural correlations resist causal interpretation. This talk explores how exponential scaling is reshaping neuroscience, showcasing successes like speech decoding and outlining the limits of current causal approaches. I argue for a shift toward tractable systems, structure–function integration, and targeted interventions. As we move from intuitive models to machine-driven discovery, we must reimagine what it means to explain the brain—not just predict it.
K2 Low Dimensional Neural Manifolds for the Control of Movement
1Department of Neuroscience, Feinberg School of Medicine, Northwestern University, Chicago, IL 60611
*Email: solla@northwestern.edu
The ability to simultaneously record the activity from tens to hundreds to thousands of neurons has allowed us to analyze the computational role of population activity as opposed to single neuron activity. Recent work on a variety of cortical areas suggests that neural function may be built on the activation of population-wide activity patterns, the neural modes, rather than on the independent modulation of individual neural activity. These neural modes, the dominant covariation patterns within the neural population, define a low dimensional neural manifold that captures most of the variance in the recorded neural activity. We refer to the time-dependent activation of the neural modes as their latent dynamics and argue that latent cortical dynamics within the manifold are the fundamental and stable building blocks of neural population activity.
A focus on motor cortex provides a model for neural control of movement in which "neural modes" are the generators of motor behaviors. We review existing evidence in support of neural manifolds, and present novel results on the relation between latent dynamics, muscle activity, and limb control. A method to align the latent dynamics associated with a simple task allows us to track manifold activity across time and across primates.
K3 Perturbations and decisions
1College of Physicians and Surgeons and Mortimer B. Zuckerman Mind Brain Behavior Institute, Columbia University, New York, NY 10027, USA
*Email: ken@neurotheory.columbia.edu
I'll present two stories. One is about our work on how perturbation responses derive from circuitry and how circuitry can be derived from perturbation responses. This will focus on responses in E/I networks to optogenetic stimulation of small numbers of cells. I'll introduce a new analytically solvable approach to linear neural field models and a novel nonlinear analysis. Surprisingly, we find that a spatially very narrow component of excitation, surrounded by suppression, does not require a correspondingly spatially narrow component of connectivity, but instead depends primarily on the strength of feedback inhibition, and make a number of other inferences and predictions about circuit structure. The second story analyzes population recordings in macaque area LIP during a reaction-time, perceptual decision-making task (deciding direction of dot motion; saccading to indicate direction as soon as the monkey decides, i.e. a reaction-time rather than fixed-delay task). We show that the trajectories in the neural state space (the space in which each dimension is the firing rate of one neuron) are organized by reaction time in a curved but locally 2D manifold. Analysis of the trajectories shows they divide into two qualitatively different portions: a "deliberation" phase in which neural responses are noisy both in cognitively relevant and irrelevant direction, are driven by fluctuations in sensory evidence, and do not strongly separate by choice; and a "commitment" phase in which noise is suppressed, influence of sensory evidence seems minimal as though the dynamics is more intrinsically-circuit-driven, and trajectories separate strongly by choice and proceed directly to the saccade endpoint. The deliberation phase is not seen for the fastest reaction times, and becomes more and more prominent with increasing reaction time. Surprisingly, the commitment phase can begin a long time (~500ms or more) before the saccade preparation, in contrast to previously prevailing ideas that saccade preparation immediately follows an internal decision reached by sensory evidence integration. A circuit model accounts for all the data: (1) activity bumps corresponding to the two possible saccade targets compete during the deliberation phase, and move continually towards one another due to recurrent circuit interactions while deliberation proceeds; (2) the longer the deliberation period, and thus the longer the reaction time, the further the bumps are displaced from the original target position; (3) commitment arises when one bump "wins" and grows rapidly, at its displaced location, under its own self-excitation and its mean input. Because of the bump movement, the model predicts that (1) choice selectivity of neurons depends on reaction time, with neurons representing positions toward fixation from the two targets being choice selective on slow trials while those representing positions away from fixation relative to the targets are choice selective for fast trials; (2) The longer the reaction time, the more the saccades tend to undershoot the saccade targets. Both predictions are confirmed in the data.
K4 Learning to infer transitively on a mental line in premotor cortex
1Natl. Center for Radiation Protection and Computational Physics, Istituto Superiore di Sanità, 00161 Roma, Italy
*Email: maurizio.mattia@iss.it
Understanding how the brain encodes and manipulates sequential knowledge remains a central challenge in neuroscience. In this talk, I will propose and empirically support a geometric framework in which high-dimensional neural representations organize knowledge along a “mental line” learned through experience [1]. This model explains how serial ordering and transitive inference emerge from mixed selectivity in neural populations, enabling robust performance even under noisy sensory and neural conditions. Integrating behavioral, computational, and neural data, I will show that premotor cortex (PMC) in monkeys encodes this mental line as animals acquire novel item relationships [2]. Neural population activity in PMC dynamically aligns with theoretical predictions, revealing a learning-dependent rotation of the “geometric mental line” that links abstract serial reasoning with motor planning. These findings highlight a mechanistic bridge between symbolic knowledge, neural geometry, and behavior, suggesting a pivotal role for motor-related associative cortices in the flexible encoding of sequential and inferential knowledge.
[1] Di Antonio, G., Raglio, S., & Mattia, M. (2024). A geometrical solution underlies general neural principle for serial ordering. Nature Communications, 15, 8238. https://doi.org/10.1038/s41467-024-52240-6
[2] Raglio, S., Di Antonio, G., Brunamonti, E., Ferraina, S., & Mattia, M. (2024). Learning to infer transitively: serial ordering on a mental line in premotor cortex. bioRxiv, 2024.10.29.620924. https://doi.org/10.1101/2024.10.29.620924
FO1 Hearing Music: A Shared Geometry Governs the Trade-off Between Reliability and Complexity in the Neural Code
Pauline G. Mouawad*1, Shievanie Sabesan1, Alinka E. Greasley2, and Nicholas A. Lesica1
1The Ear Institute, University College London, London, UK
2School of Music, University of Leeds, Leeds, UK
*Email: p.mouawad@ucl.ac.uk
Music is central to human culture, shaping social bonds and emotional well-being. Its unique ability to connect sensory processing with reward, emotion, and statistical learning makes it an ideal tool for studying auditory perception [1]. Previous studies have explored neural responses to speech and to simple musical sounds [2, 3], but the neural coding of complex music remains unexplored. We addressed this gap by analyzing multi-unit activity (MUA) recorded from the inferior colliculus (IC) of normal-hearing (NH) and hearing-impaired (HI) gerbils in response to a range of music types at multiple sound levels. The music types included individual stems (vocals, drums, bass, and other) as well as mixtures in which the stems were combined.
Using coherence analysis, we assessed how reliably music is encoded in the IC across repeated presentations of stimuli and the degree to which individual stems are distorted when presented in a mixture. To explore neural activity patterns at the network level, we implemented a manifold analysis using PCA. This identified the signal manifold, the subspace where reliable musical information is embedded. To model neural transformations underlying music encoding, we developed a deep neural network (DNN) capable of generating MUA from sound, providing a framework for interpreting how the IC processes music. Finally, to assess the impact of hearing loss, we conducted a comparative analysis for NH and HI at equal sound and sensation levels.
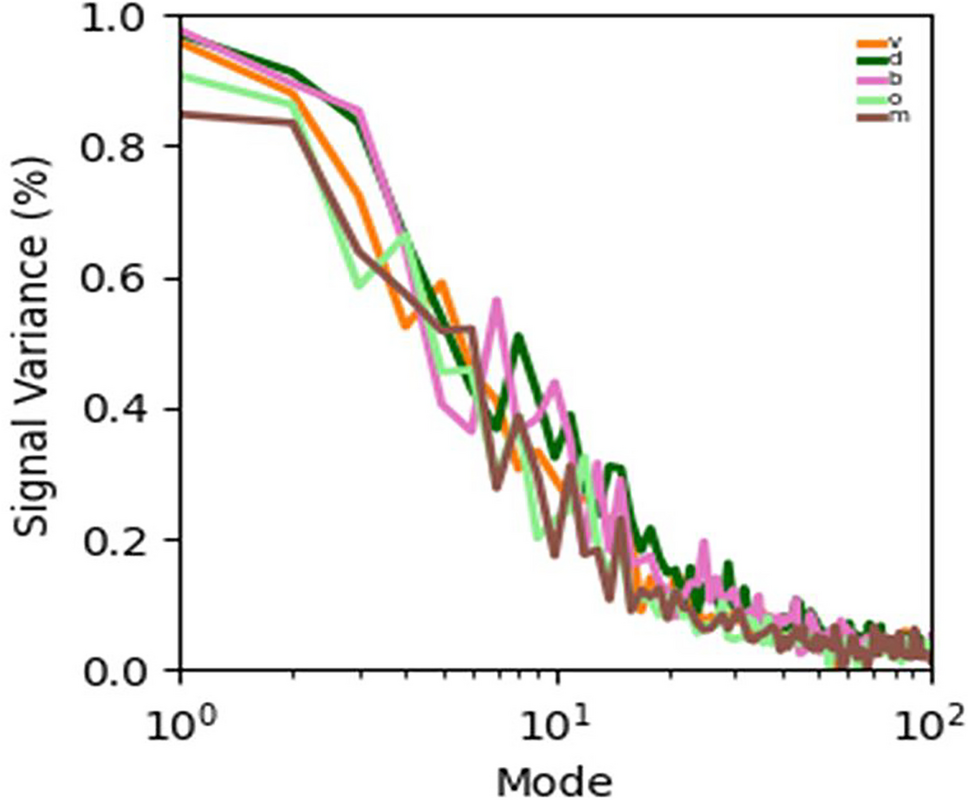
We identified strong nonlinear interactions between stems, affecting both the reliability and geometry of neural coding. The reliability of the responses and the dimensionality of the signal manifold varied widely across music types. With increasing musical complexity, the dimensionality of the signal manifold increased, however the reliability decreased. The leading modes in the signal manifold were reliable and shared across all music types, but as musical complexity increased, new neural modes emerged, though these were increasingly unreliable (Figure 1). Our DNN successfully synthesized MUA from music with high fidelity. After hearing loss, neural coding was strongly distorted at equal sound level, but these distortions were largely corrected at equal sensation level.
Music processing in the early auditory pathway involves nonlinear interactions that shape the neural representation in complex ways. The signal manifold contains a fixed set of leading modes that are invariant across music types. As music becomes more complex the manifold is not reconfigured; instead, new, less reliable modes are added. These new modes reflect a fundamental trade-off between fidelity and complexity in the neural code. The fact that suitable amplification restores near-normal neural coding suggests that mild-to-moderate hearing loss primarily affects audibility rather than the brainstem’s capacity to process music.
Acknowledgment Funding for this work was provided by the UK Medical Research Council through grant MR/W019787/1.
[1] Juslin, P. & Västfjäll, D. (2008). Emotional responses to music: The need to consider underlying mechanisms. Behavioral and Brain Sciences, 31(5), 559-575. doi: 10.1017/S0140525X08005293
[2] Rajendran, V., Harper, N., Garcia-Lazaro, J., Lesica, N., & Schnupp, J. (2017). Midbrain adaptation may set the stage for the perception of musical beat. Proceedings of the Royal Society B: Biological Sciences, 284(1866), 20171455. doi: 10.1098/rspb.2017.1455
[3] Sabesan, S., Fragner, A., Bench, C., Drakopoulos, F., & Lesica, N. A. (2023). Large-scale electrophysiology and deep learning reveal distorted neural signal dynamics after hearing loss. Elife, 12, e85108.

Figure 1: Complexity and Reliability in the Latent Space
FO2 Global brain dynamics modulates local scale-free neuronal activity
Giovanni Rabuffo*1,2, Pietro Bozzo1, Marco Pompili1, Damien Depannemeacker1, Bach Nguyen2, Tomoki Fukai2, Pierpaolo Sorrentino1, and Leonardo Dalla Porta3
1Institut de Neurosciences des Systèmes (INS), Aix Marseille University, Marseille, France
2Okinawa Institute for Science and Technology (OIST), Okinawa, Japan
3Institute of Biomedical Investigations August Pi i Sunyer (IDIBAPS), Systems Neuroscience, Barcelona, Spain
*Email: giovanni.rabuffo@univ-amu.fr
The brain's ability to balance stability and flexibility is thought to emerge from operating near a critical state [1]. In this work we address two major gaps of the “brain criticality hypothesis”: First, local (between neurons) and global (between brain regions) criticality are often investigated independently, and a unifying framework is lacking. Second, local neuronal populations do not maintain a strictly critical state but rather fluctuate around it [2]. The mechanisms underlying these fluctuations remain unclear. To bridge these gaps, we introduce a connectome-based model that allows for a simultaneous assessment of local and global criticality (Fig.1). We demonstrate that long-range structural connectivity shapes global critical dynamics and drives the fluctuations of each brain region around a local critical state.
Decoupled brain regions are described by a mean-field model [3] which exhibits avalanche-like dynamics under stochastic input (Fig.1, Blue). Brain regions are connected via the Allen Mouse Connectome [4], and simulations are performed for different values of the global coupling parameter [5]. Simulated data consists of fast LFP, and slow BOLD signals (Fig.1, Red). The model results are validated against empirical datasets (Fig.1, Gray), including a mouse fMRI dataset [6] and LFP recordings from the Allen Neuropixel dataset [7]. To quantify the fluctuations around criticality, we identified neuronal avalanches as deviations of the local LFP signals below a fixed threshold (Fig.1, Blue) and measured sizes (area under curve) and durations (time to return within threshold). The magnitude of the fluctuations around criticality is assessed by analyzing the variance of the range of avalanche sizes across 2s-long epochs.
For low global coupling, individual brain regions maintains local criticality (Fig.1, Blue) but remains globally desynchronized. Increasing coupling induces spontaneous long-range synchronization, paralleled by local fluctuations around criticality (Fig.1, Red). Notably, the working point where the simulations match the experiments corresponds to the regime with the largest range of avalanches sizes and durations (Fig.1, Grey). Strongly connected regions exhibit greater fluctuations around criticality, a testable prediction of the model. To verify this, we examined Allen Mouse Brain Atlas ROIs with LFP data and found a significant correlation between empirical critical fluctuations and regional structural connectivity properties (Fig.1, Green).
Our results, comparing brain simulations and empirical datasets across scales, support the brain criticality hypothesis and suggest that criticality is not a static regime for a local neuronal population, but it is dynamically up- and down- regulated by large-scale interactions.
Acknowledgment We thank the Institut de Neurosciences des Systèmes (INS), Marseille, France, and the Okinawa Institute for Science and Technology, Japan for their generous support and sponsorship of this research. Their contributions have been instrumental in advancing our understanding of brain criticality and its implications.
[1] O’Byrne, J. & Jerbi, K. (2022). How critical is brain criticality?. Trends in Neurosciences, 45(11), 820-837. doi: 10.1016/j.tins.2022.08.007
[2] Fontenele, A., de Vasconcelos, N., Feliciano, T., Aguiar, L., Soares-Cunha, C., Coimbra, B., Dalla Porta, L., Ribeiro, S., Rodrigues, A., Sousa, N., Carelli, P., & Copelli, M. (2019). Criticality between Cortical States. Physical Review Letters, 122(20). doi: 10.1103/physrevlett.122.208101
[3] Buendía, V., Villegas, P., Burioni, R., & Muñoz, M. (2021). Hybrid-type synchronization transitions: Where incipient oscillations, scale-free avalanches, and bistability live together. Physical Review Research, 3(2). doi: 10.1103/physrevresearch.3.023224
[4] Oh, S., Harris, J., Ng, L., Winslow, B., et al. (2014). A mesoscale connectome of the mouse brain. Nature, 508(7495), 207-214. doi: 10.1038/nature13186
[5] Melozzi, F., Woodman, M., Jirsa, V., & Bernard, C. (2017). The Virtual Mouse Brain: A Computational Neuroinformatics Platform to Study Whole Mouse Brain Dynamics. eneuro, 4(3), ENEURO.0111-17.2017. doi: 10.1523/eneuro.0111-17.2017
[6] Grandjean, J., Desrosiers-Gregoire, G., Anckaerts, C., et al. (2023). A consensus protocol for functional connectivity analysis in the rat brain. Nature Neuroscience, 26(4), 673-681. doi: 10.1038/s41593-023-01286-8
[7] Allen Institute for Brain Science. (n.d.). Visual coding with Neuropixels. Allen Institute for Brain Science. https://allensdk.readthedocs.io/en/latest/visual_coding_neuropixels.html

Figure 1: (Blue) Local neural mass model displays critical-like avalanche dynamics. (Red) Coupling brain regions via the empirical Allen structural connectivity we simulate fast LFP and slow BOLD global dynamics. (Gray) Simulated LFP displays global critical activity and simulated BOLD data matches fMRI experiments. (Green) The fluctuations around criticality correlate with structural in-strength.
FO3 Single-cell optogenetic perturbations reveal stimulus-dependent network interactions
Deyue Kong*1, Joe Barreto2, Greg Bond2, Matthias Kaschube1, and Benjamin Scholl2
1Frankfurt Institute for Advanced Studies, Frankfurt am Main, Germany
2University of Colorado Anschutz Medical Campus, Department of Physiology and Biophysics, Aurora, Colorado, USA
*Email: kong@fias.uni-frankfurt.de
Cortical computations arise through neuronal interactions and their dynamic reconfiguration in response to changing sensory contexts. Cortical interactions are proposed to engage distinct operational regimes that either amplify or suppress particular neuronal networks. A recent study in mouse primary visual cortex (V1) found competitive, suppressive interactions between nearby, similarly-tuned neurons, with exception of highly-correlated neuronal pairs showing facilitatory coupling [1]. It remains unclear whether such feature competition generalizes to cortical circuits with topographic organization, where neighboring neurons within columns exhibit similar tuning to visual features, and distal excitatory axons preferentially target similarly-tuned columns.
We investigated interactions between excitatory neurons in the ferret V1 and how network interactions depend on stimulus strength (contrast). We recorded the responses of layer 2/3 neurons to drifting gratings of eight directions at two contrast levels using 2-photon calcium imaging, while activating individual excitatory neurons with precise 2-photon optogenetics. We statistically quantified the effect of target photostimulation on neural activity (inferred spike rate) during visual stimulation using a Poisson generalized linear model (GLM). We then used our model to estimate a target’s influence on the surrounding neurons’ activity and their stimulus coding properties.
Our analyses revealed interactions that depended on cortical distance, stimulus properties, and functional similarity between neuron pairs. Influence of photostimulated neurons strongly depended on cortical distance, but overall exhibited net suppression. Suppression was weakest between nearby neurons (<100μm), but was found across large cortical distances. Distant-dependent suppression was reduced when visual stimuli were low contrast. Examining functional-similar neurons, we found that noise correlations between neuron pairs were most predictive of measured interactions, showing a strong shift from amplification to competition: at low contrast, we observed local amplification between noise-correlated excitatory neurons, but increasing contrast led to a predominantly suppressive influence across all distances.
Our data support predictions from theoretical models, such as stabilized supralinear networks (SSN), in which networks amplify weak feed-forward input, but sublinearly integrate strong inputs [2,3]. Furthermore, decoding analyses suggest that the contrast-dependent shift from facilitation to suppression correlates with improved decoding accuracy of direction. These findings demonstrate that stimulus contrast dynamically modulates recurrent interactions between excitatory neurons in ferret V1, likely by differentially engaging inhibitory neurons. Such dynamic modulation supports optimal encoding of sensory information within columnar cortices.
[1] Chettih, S. & Harvey, C. (2019). Single-neuron perturbations reveal feature-specific competition in V1. Nature, 567(7748), 334-340. doi: 10.1038/s41586-019-0997-6
[2] Rubin, D. B., Van Hooser, S. D., & Miller, K. D. (2015). The stabilized supralinear network: a unifying circuit motif underlying multi-input integration in sensory cortex. Neuron, 85(2), 402-417.
[3] Heeger, D. J., & Zemlianova, K. O. (2020). A recurrent circuit implements normalization, simulating the dynamics of V1 activity. Proceedings of the National Academy of Sciences, 117(36), 22494-22505.
FO4 Automated identification of disease mechanisms in hiPSC-derived neuronal networks using simulation-based inference
Nina Doorn*1, Michel van Putten1,2, and Monica Frega3
1Department of Clinical Neurophysiology, University of Twente, Enschede, The Netherlands
2Department of Neurology and Clinical Neurophysiology, Medisch Spectrum Twente, The Netherlands
3Department of Informatics, Bioengineering, Robotics and System Engineering, University of Genova, Italy
*Email: n.doorn-1@utwente.nl
Human induced pluripotent stem cells (hiPSCs)-derived neuronal networks on multi-electrode arrays (MEAs) are a powerful tool to study neurological disorders in vitro [1]. The electric activity patterns of these networks differ between healthy and patient-derived neurons, reflecting underlying pathology (Fig. 1A). However, elucidating the underlying molecular mechanisms is challenging and requires extensive, costly, and hypothesis-driven additional experiments. Biophysical models can link observable network activity to underlying molecular mechanisms by estimating model parameters that simulate the experimental observations. However, parameter estimation in such models is difficult due to stochasticity, non-linearity, and parameter degeneracy.
Here, we address this challenge using simulation-based inference (SBI), a machine-learning approach that allows efficient statistical inference of biophysical model parameters using only simulations [2]. We apply SBI to our previously validated biophysical model of hiPSC-derived neuronal networks on MEA [3], which includes Hodgkin-Huxley-type neurons and detailed synaptic models (Fig. 1B). To train SBI, we simulated 300,000 network configurations, varying key parameters governing synaptic and intrinsic neuronal properties (Fig. 1C)[4]. We used a neural density estimator to infer posterior distributions of these model parameters given experimental MEA recordings from healthy, pharmacologically treated, and patient-derived networks (Fig 1D)[4].
SBI accurately inferred ground-truth parameters in synthetic data and successfully identified known disease mechanisms in patient-derived neuronal networks. In networks from patients with the genetic epilepsies Dravet Syndrome and GEFS+, SBI predicted reduced sodium and potassium conductances and increased synaptic depression, which was experimentally verified. In CACNA1A haploinsufficient networks, SBI correctly identified impaired connectivity. Additionally, SBI detected drug-induced changes, such as prolonged synaptic depression following Dynasore treatment.
SBI enables automated and probabilistic inference of biophysical parameters, offering advantages over traditional parameter estimation methods, which can be time-consuming, lack uncertainty quantification, or cannot deal with parameter degeneracy. Our results show how SBI can be used with biophysical models to identify possible disease mechanisms from patient-derived neuronal data. Our proposed analysis pipeline enables researchers to extract crucial mechanistic information from MEA measurements in a systematic, cost-effective, and rapid manner, paving the way for targeted experiments and novel insights into disease.
Acknowledgment This work was supported by the Netherlands Organisation for Health Research and Development ZonMW; BRAINMODEL PSIDER program 10250022110003 (to M.F.). We thank Eline van Hugte, Marina Hommersom, and Nael Nadif Kasri for providing MEA recordings from patient-derived and genome-edited in vitro neuronal networks.
[1] Mossink, B., Verboven, A., van Hugte, E., Klein Gunnewiek, T., Parodi, G., Linda, K., Schoenmaker, C., Kleefstra, T., Kozicz, T., van Bokhoven, H., Schubert, D., Nadif Kasri, N., & Frega, M. (2021). Human neuronal networks on micro-electrode arrays are a highly robust tool to study disease-specific genotype-phenotype correlations in vitro. Stem Cell Reports, 16(9), 2182-2196. doi: 10.1016/J.STEMCR.2021.07.001
[2] Gonçalves, P., Lueckmann, J., Deistler, M., Nonnenmacher, M., Öcal, K., Bassetto, G., Chintaluri, C., Podlaski, W., Haddad, S., Vogels, T., Greenberg, D., & Macke, J. (2020). Training deep neural density estimators to identify mechanistic models of neural dynamics. eLife, 9. doi: 10.7554/ELIFE.56261
[3] Doorn, N., Voogd, E., Levers, M., van Putten, M., & Frega, M. (2024). Breaking the burst: Unveiling mechanisms behind fragmented network bursts in patient-derived neurons. Stem Cell Reports, 19(11), 1583-1597. doi: 10.1016/J.STEMCR.2024.09.001
[4] Doorn, N., van Putten, M. J., & Frega, M. (2025). Automated inference of disease mechanisms in patient-hiPSC-derived neuronal networks. Communications Biology, 8(1), 768.

Figure 1: A) The activity of in vitro neuronal networks cultured from hiPSCs of healthy controls and patients is measured using MEAs. B) The computational model with biophysical parameters in blue. C) A Neural density estimator is trained on model simulations. Afterward, experimental data is passed through the estimator to approximate the D) posterior distributions. Adapted from [4].
O1 Balancing Stability and Flexibility: Dynamical Signatures of Learning in In-Vitro Neuronal Networks
Forough Habibollahi*1, and Brett J. Kagan1
1Cortical Labs, Melbourne, Australia
*Email: forough@corticallabs.com
CL1 is a novel system which bridges biological intelligence and adaptive neuronal traits by integrating in-vitro neuronal networks with in-silico computational elements using micro-electrode arrays (MEAs) [1]. These cultivated neuronal ensembles demonstrate self-organized, biological adaptive intelligence in dynamic gaming environments via closed-loop stimulation and concurrent recordings. While in-vitro neuronal networks are shown to achieve real-time adaptive learning, the underlying network dynamics enabling this learning remain under explored.
We investigated pairwise causal relationships between recorded channels using Granger causality analysis [2], reconstructing a connectivity network from statistically significant causal interactions. The most influential/influenced nodes were identified as those with highest outgoing/incoming connections. To explore dynamic properties, we reconstructed the phase space of the spiking time series from all recorded channels using state-space reconstruction [3]. Optimal embedding dimensions were determined by minimizing false nearest neighbors, while time delays were selected by detecting the first local minimum of mutual information across different delays. Recurrence plots were generated from the reconstructed phase spaces to analyze temporal patterns.
We analyzed 45-minute spiking recordings at 25 kHz from 23 neuronal cultures, comprising 111 rest sessions and 133 gameplay sessions. Across both rest and gameplay conditions, we observed distinct dynamic patterns between “influential” and “influenced” nodes. Overall, the gameplay sessions exhibited higher recurrence (RR) and determinism (DET) compared to rest (Fig. 1.a). However, in both conditions, the “influenced” nodes displayed lower RR and more negative Lyapunov exponents—indicative of more ordered behavior that lies farther from the edge of chaos. In contrast, the most influential nodes showed higher RR, reflecting recurrent and cyclic dynamics, and had small negative Lyapunov exponents, consistent with behavior near the edge of chaos (Fig 1.b.).
Our findings reveal a functional dichotomy in in-vitro neuronal networks. Influential channels exhibit cyclic behavior near the edge of chaos, marked by high RR and near-zero negative Lyapunov exponents, balancing order and chaos. These “near-chaotic” nodes drive network dynamics, enabling rapid influence and adaptability. In contrast, influenced channels remain more ordered, with lower recurrence and more negative Lyapunov exponents, suggesting stable responsiveness. This interplay between near-chaotic drivers and stable receivers enables neuronal cultures to balance robustness with adaptability. By defining how distinct dynamical states interact, our results shed light on coordinated neuronal activity and the role of near-chaotic dynamics in flexible behavior.
Acknowledgment F.H. and B.J.K. are employees of Cortical Labs. B.J.K. is a shareholder of Cortical Labs. B.J.K. holds an interest in patents related to this publication.
[1] Kagan, B., Kitchen, A., Tran, N., Habibollahi, F., Khajehnejad, M., Parker, B., Bhat, A., Rollo, B., Razi, A., & Friston, K. (2022). In vitro neurons learn and exhibit sentience when embodied in a simulated game-world. Neuron, 110(23), 3952-3969.e8. doi: 10.1016/j.neuron.2022.09.001
[2] Granger, C. (1969). Investigating Causal Relations by Econometric Models and Cross-spectral Methods. Econometrica, 37(3), 424. doi: 10.2307/1912791
[3] Takens, F. (1981). Detecting strange attractors in turbulence. Lecture Notes in Mathematics, 366-381. doi: 10.1007/BFb0091924

Figure 1: a) Comparison of dynamic metrics between rest and gameplay sessions. Bar plots show mean values (±SEM) for recurrence rate (RR), determinism (DET), laminarity (LAM), and Lyapunov exponent across all recorded electrodes. b) Dynamic properties of influential vs. influenced nodes across rest and gameplay conditions.
O2 Representational drift as a correlate of memory consolidation
Denis Alevi*+,1,2, Felix Lundt+,1, Simone Ciceri1, Kristine Heiney1, and Henning Sprekeler1,2,3
1Modelling of Cognitive Processes, Technische Universität Berlin, Berlin, Germany
2Bernstein Center for Computational Neuroscience Berlin, Berlin, Germany
3Science of Intelligence, Research Cluster of Excellence, Berlin, Germany
*Email: denis.alevi@tu-berlin.de
Neural representations – and their population geometry – often change over time despite stable behavior, a phenomenon termed representational drift [1-4]. It is debated if drift is driven by a random process or if it has a directed component, and if it serves a computational function [5]. Systems memory consolidation is a promising candidate [6], because it predicts a temporal reorganization of neural memory engrams. However, it remains unclear how classical theories of consolidation relate to the population-level view of drift and how apparently unstructured drift could be driven by a directed consolidation process.
We present a computational model for engram dynamics under memory consolidation and explore the resulting representational drift. Assuming that engram changes are driven by reactivations, the model displays recurrent neural network (RNN)-like dynamics, but evolves on long time scales of memory consolidation. This allows us to reinterpret common dynamical phenomena in RNNs in light of memory consolidation and relate them to experimentally observed drift. In simulation, we study how single cell tuning curves and the geometry of neural representations change over time, when not all neurons are observed and develop analytical results for the effect of subsampling, based on Green’s functions and random matrix theory.
Our model redistributes memory engrams across neural populations while maintaining stable memory recall through null-space dynamics [2]. The model can display power-law forgetting without requiring a diversity of learning rates [7]. Low-rank dynamics induce selective consolidation and semantization. In line with experimental findings on representational drift, individual neurons exhibit diverse tuning changes: stability, gradual drift, and abrupt changes of preferred stimulus. Multi-day decoders [2] reveal invariant subspaces on the full population, but degrade quickly under subsampling. A theoretical analysis shows that the dynamics of subsampled populations can be predominantly driven by the unrecorded population, which generates seemingly noise-driven dynamics.
Our phenomenological model of engram dynamics bridges the gap between the area-centered perspective of systems consolidation and the population-level perspective of representational drift. Our results show that despite systematic population dynamics, a recorded subset of the neural population can appear to have unstructured dynamics [2]. Recent evidence for stable geometric structure during representational drift in CA1 [7] is consistent with our model of RNN-like engram dynamics, and we hypothesize that unstable population geometry [3] could also be explained by subsampling. Overall, our model offers a functional interpretation of drift as a means to redistribute engrams for improved memory retention.
Acknowledgment Kristine Heiney is funded by a Postdoctoral Research Fellowship from the Alexander von Humboldt Foundation.
[1] Driscoll, L., Pettit, N., Minderer, M., Chettih, S., & Harvey, C. (2017). Dynamic Reorganization of Neuronal Activity Patterns in Parietal Cortex. Cell, 170(5), 986-999.e16. doi: 10.1016/j.cell.2017.07.021
[2] Rule, M., Loback, A., Raman, D., Driscoll, L., Harvey, C., & O'Leary, T. (2020). Stable task information from an unstable neural population. eLife, 9. doi: 10.7554/eLife.51121
[3] Schoonover, C., Ohashi, S., Axel, R., & Fink, A. (2021). Representational drift in primary olfactory cortex. Nature, 594(7864), 541-546. doi: 10.1038/s41586-021-03628-7
[4] Masset, P., Qin, S., & Zavatone-Veth, J. (2022). Drifting neuronal representations: Bug or feature?. Biological Cybernetics, 116(3), 253-266. doi: 10.1007/s00422-021-00916-3
[5] Driscoll, L., Duncker, L., & Harvey, C. (2022). Representational drift: Emerging theories for continual learning and experimental future directions. Current Opinion in Neurobiology, 76, 102609. doi: 10.1016/j.conb.2022.102609
[6] Roxin, A. & Fusi, S. (2013). Efficient Partitioning of Memory Systems and Its Importance for Memory Consolidation. PLoS Computational Biology, 9(7), e1003146. doi: 10.1371/journal.pcbi.1003146
[7] Sylte, O. C., Kilias, A., Bartos, M., & Sauer, J. F. (2025). Coordinated representational drift supports stable place coding in hippocampal CA1. bioRxiv, 2025-02.
O3 Backpropagation through space, time and the brain
Paul Haider*1, Benjamin Ellenberger1, Jakob Jordan1, Kevin Max1, Ismael Jaras1, Laura Kriener1, Federico Benitez1, and Mihai A. Petrovici1
1Department of Physiology, University of Bern, Bern, Switzerland
*Email: paul.haider@unibe.ch
Effective learning in the brain relies on the adaptation of individual synapses based on their relative contribution to solving a task. However, the challenge of spatio-temporal credit assignment in physical neuronal networks remains largely unsolved due to the biologically implausible assumptions of traditional backpropagation algorithms. This study aims to bridge this gap by proposing a novel framework that efficiently performs credit assignment in real-time, without violating spatio-temporal locality constraints, driven by the need for biological systems to learn continuously and interact with dynamic environments.
We introduce Generalized Latent Equilibrium (GLE), a computational framework for fully local spatio-temporal credit assignment in physical, dynamical networks of neurons. GLE is based on an energy function of neuron-local mismatch errors, from which neuronal dynamics are derived using stationarity and parameter dynamics using gradient descent principles. This framework leverages the morphology of dendritic trees and the ability of neurons to phase-shift their output rates relative to their input (see, e.g., [1]), enabling complex information processing. Additionally, the adjoint method is employed to demonstrate that our learning rules approximate gradient descent on the total integrated cost over time, effectively approximating backpropagation through time (BPTT).
The resulting neuronal dynamics can be interpreted as a real-time, biologically plausible approximation of backpropagation through space and time, incorporating continuous-time leaky-integrator neuronal dynamics and continuously active, phase-free, local synaptic plasticity. The corresponding equations suggest a direct mapping to cortical microcircuitry, with L2/3 pyramidal error neurons counter-posing L5/6 pyramidal representation neurons in a ladder-like fashion. We demonstrate GLE's effectiveness on both spatial and temporal tasks, such as chaotic time series prediction, MNIST-1D [2], and Google Speech Commands datasets, achieving results competitive with powerful ML architectures like GRUs and TCNs trained with offline BPTT.
This framework has significant implications for understanding biological learning processes in neural circuits and designing neuromorphic hardware. GLE is applicable to both spatial and temporal tasks, offering advantages over existing alternatives like BPTT and real-time recurrent learning (RTRL) in terms of efficiency and biological plausibility. The framework's locality and reliance on conventional analog components make it an attractive blueprint for efficient neuromorphic hardware. This study contributes to a deeper understanding of how physical neuronal systems can efficiently learn and process information in real-time, bridging the gap between machine learning and biological neural networks.
Acknowledgment This work was supported by the European Union, the Volkswagen Foundation, ESKAS, and the Manfred Stärk Foundation. We also acknowledge the Fenix Infrastructure and the Insel Data Science Center for their support.
[1] Brandt, S., Petrovici, M. A., Senn, W., Wilmes, K. A., & Benitez, F. (2024). Prospective and retrospective coding in cortical neurons. arXiv preprint arXiv:2405.14810.
[2] Greydanus, S., & Kobak, D. (2020). Scaling down deep learning with mnist-1d. arXiv preprint arXiv:2011.14439.
O4 Competition between memories for reactivation as a mechanism for long-delay credit assignment
Subhadra Mokashe*1, and Paul Miller2
1Neuroscience Graduate Program, Brandeis University, Waltham, USA
2Department of Biology, Brandeis University, Waltham, USA
*Email: subhadram@brandeis.edu
Animals learn to associate an event with its outcome, as in conditioned taste aversion, when they gain aversion to a conditioned stimulus (CS, recently experienced taste) if sickness is later induced [1]. Overshadowing arises if another intervening taste (interfering stimulus, IS) gains some credit for the causality of the outcome, thereby reducing the aversion to the CS [2]. The known short-term correlational plasticity mechanisms do not wholly explain how networks of neurons achieve long-delay credit assignment. We hypothesize that reactivation of stimuli during sickness causes specific associative learning between those stimuli and the sickness, and the competition between the stimuli for reactivation could explain overshadowing.
We build a spiking recurrent network model with clustered connectivity for excitatory neurons and unstructured inhibitory feedback. We assume the recurrent strengths are enhanced at the time of stimulus presentation due to Hebbian mechanisms and then decay in time. Given that the IS is introduced after the CS, the IS ensemble has higher recurrent strength than the CS ensemble. When we simulate the network, we see reactivation of both tastes (Fig 1 A). We calculate the fraction of time the network spends reactivating a stimulus as a readout of association with the outcome (sickness). We vary the interstimulus interval by changing the difference in recurrent strengths (Δ) and vary the delay to sickness by varying the recurrent strengths.
When we look at the time spent in each state as we increase Δ, we see that not only the time spent in the IS increases, but the time spent in the CS decreases (Fig. 1 B). We only changed the recurrent strengths of the IS ensemble; the time spent in the CS ensemble was affected, indicating competition between the memories for reactivation and accounts for overshadowing. When the CS to IS interval is held constant, paradoxically, more conditioning to the CS is shown by a later sickness onset than earlier sickness [2]. We can explain the result via greater time spent in the CS state (Fig. 1 D) with an appropriate decay profile of recurrent weights (Fig. 1 C) such that the reduced overshadowing outweighs the reduction in conditioning with increased delay.
How actions are associated with delayed outcomes is not well understood. We explore the reactivation of memories as a mechanism for long-delay credit assignment in conditioned taste aversion (CTA). We show that competition between memories for reactivation could explain how credit is assigned when there is ambiguity about the cause of an outcome. We use theoretical predictions to constrain our model and are able to explain experimental findings for overshadowing [2]. This study could explain credit assignment not only in CTA and overshadowing but also in other forms of long-delay learning and provide insights into how credit is assigned when there is ambiguity in the cause of an outcome.
Acknowledgment We acknowledge Donald Katz and Hannah Germaine for discussions about the work. We thank NIH, NINDS for funding via R01 NS104818.
[1] Nachman, M. (1970). Learned taste and temperature aversions due to lithium chloride sickness after temporal delays.. Journal of Comparative and Physiological Psychology, 73(1), 22-30. doi: 10.1037/h0029807
[2] Kwok, D., Harris, J., & Boakes, R. (2016). Timing of interfering events in one-trial serial overshadowing of a taste aversion. Learning & Behavior, 45(2), 124-134. doi: 10.3758/s13420-016-0246-x

Figure 1: A. Reactivation of the stimuli. B. Fraction of time spent by the network in stimuli states as a function of Δ. C. Time spent in the CS state as a function of the recurrent strength Δ, specific decay profile of the recurrent weights (red line). D. Rebound seen in the time spent in the CS state as a function of delay to the sickness onset only in the presence of the IS (red line).
O5 Acetylcholine Waves and Dopamine Release in the Striatum: A Reaction-Diffusion Mechanism
Lior Matityahu1, Naomi Gilin1, Gideon A. Sarpong2, Yara Atamna1, Lior Tiroshi1, Nicolas X. Tritsch3, Jeffery R. Wickens2, and Joshua A. Goldberg*1
1Department of Medical Neurobiology, Institute of Medical Research Israel-Canada, The Faculty of Medicine, The Hebrew University of Jerusalem, Jerusalem, Israel
2Okinawa Institute of Science and Technology Graduate University, Okinawa, Japan
3Neuroscience Institute, New York University Grossman School of Medicine, New York, NY, USA
*Email: joshua.goldberg2@mail.huji.ac.il
Striatal dopamine (DA) encodes reward and exhibits traveling waves across the mediolateral axis during behavior. However, the mechanism generating these patterns remains unknown. Cholinergic interneurons (CINs) modulate DA release through nicotinic acetylcholine receptors (nAChRs) on DA terminals. We hypothesized that reciprocal interactions between CINs and DA axons might underlie wave generation. Here, we investigated whether acetylcholine (ACh) exhibits wave-like activity, whether nAChRs extend DA release spatial scale, and whether a reaction-diffusion framework can explain these waves' emergence from local interactions.
We imaged ACh sensors (GRAB-ACh3.0, iAChSnFR) in the dorsal striatum of head-fixed mice through cranial windows and GRIN lenses. To test whether nAChRs extend DA release, we expressed GRAB-DA2m in striatal DA axons and measured electrically-evoked DA release at increasing distances with and without the nAChR antagonist mecamylamine. We combined patch-clamp recordings of individual CINs with two-photon imaging of GRAB-DA2m to test if single CINs trigger DA release. We developed and analyzed activator-inhibitor reaction-diffusion models of CIN-DA interactions, exploring how parameters influence wave behavior.
We observed ACh waves propagating primarily lateral-to-medial at velocities of ±10 mm/s. Mecamylamine reduced DA release spatial scale by approximately 50% (from ~532 μm to ~264 μm). Action potentials in individual CINs induced local DA release. We will present novel in vivo data showing that chemogenetic silencing of CINs reduces the spatial scale of ongoing DA release events in awake mice, directly confirming CINs' role in extending DA release. Our modeling demonstrated that CIN-DA interactions form an activator-inhibitor system generating traveling waves. Phase-nullcline-flow analysis (Fig. 1) revealed that wave properties depend on system parameters, explaining directional biases in behavioral contexts.
Our findings provide evidence for striatal ACh waves and establish that local CIN-DA fiber interactions drive endogenous traveling waves. The new in vivo data showing CINs extend DA release validates our model's core assumption. The reaction-diffusion framework explains how waves emerge from local axo-axonal interactions without external pacemakers. Our model predicts: strongly coupled DA-ACh waves, nAChR blockade compromising wave propagation, and interneuron activity influencing wave direction. This mechanism contributes to spatiotemporal coding in the striatum, with implications for reward processing, learning, and movement coordination.
Acknowledgment This work was funded by a Research Grant from the Human Frontier Science Program (RGP0062/2019), an ERC Consolidator Grant (646886), and grants from the National Institutes of Health (DP2NS105553 and R01MH130658) and Dana and Whitehall Foundations.

Figure 1: Phase-nullcline-flow analysis of the activator-inhibitor model. (a) Nullclines and flow field showing fixed points. (b) The direction of wave propagation depends on the area between nullclines. β values control the coupling strength between CINs and DA axons, determining whether CIN waves advance (β=1.0) or recede (β=1.8).
O6 Mathematical insights into the spatial heterogeneity of extracellular serotonin induced by the geometry and dynamics of serotonergic fibers
Merlin Pelz*1, Skirmantas Janusonis2, and Gregory Handy1,2
1School of Mathematics, University of Minnesota, Minneapolis, USA
2Department of Psychological and Brain Sciences, University of California, Santa Barbara, USA
All vertebrate brains, from fish to humans, contain dense meshworks of axons (fibers) that release serotonin, a key signaling molecule. The role of this massive system is poorly understood, with no analogs in current AI architectures, but it appears to support neuroplasticity. Its effects on neural networks are exerted through serotonin receptors whose activation depends on serotonin molecules in the local extracellular space. Recent studies have revealed a lack of fundamental understanding of the spatiotemporal characteristics of extracellular serotonin [1]. In particular, its concentration may vary greatly within microscopic volumes and over short time frames. Such sustained heterogeneity may be a key feature of the plastic brain.
To investigate how the geometry of the spatial arrangement of release/reuptake sites (i.e., fiber varicosities [2,3]; Fig. 1(a), (b)) and the timing of release shape serotonin concentrations in microscopic brain volumes, we extend previous work [4] and consider a 2D compartmental-reaction diffusion system that is analytically tractable. Each varicosity is modeled as a small disk where the kinetics of serotonin release and uptake (adapted from [5]) are implemented. The disks interact with the surrounding diffusive space through an infinitely permeable boundary (Fig. 1(c), (d)). This system can be rigorously reduced to an integro-ordinary-differential system that can be numerically solved efficiently.
Our system highlights precise coupling terms across varicosities that capture the diffusive memory dependence and global coupling and can be solved using arbitrary serotonin reaction kinetics at the varicosities. Using biologically realistic parameters, we observe that the serotonin concentration exhibits large temporal and spatial va
Comments (0)