
Remember me
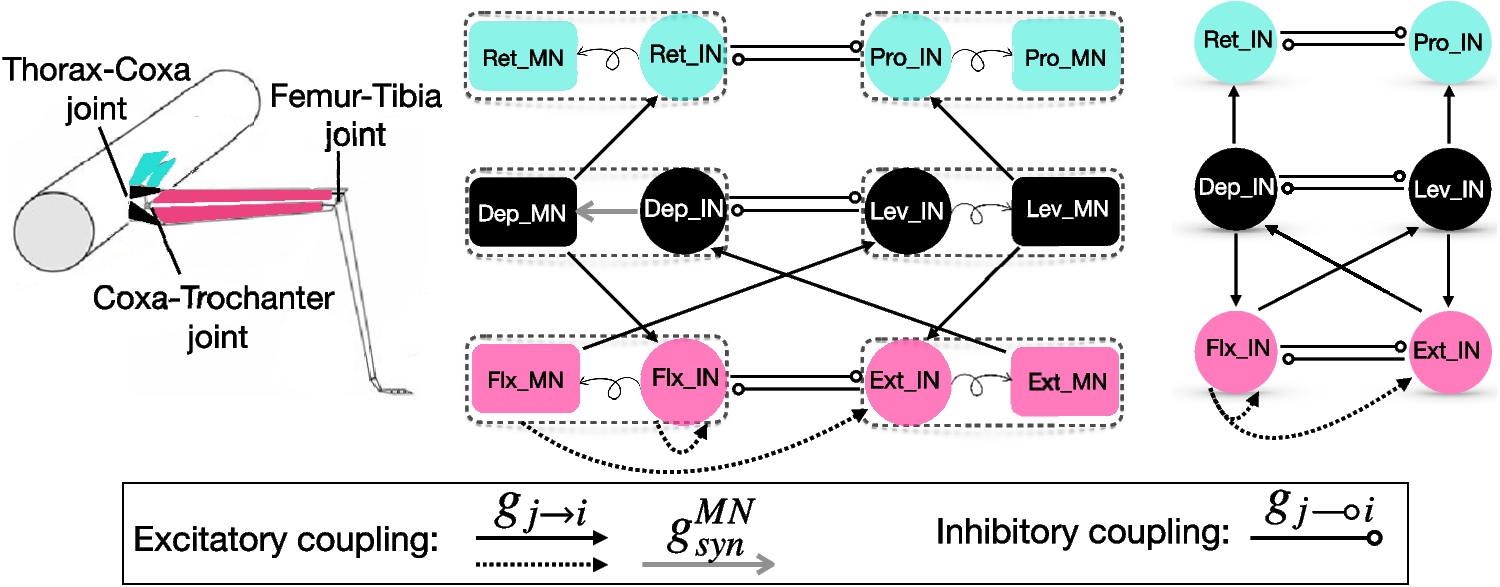
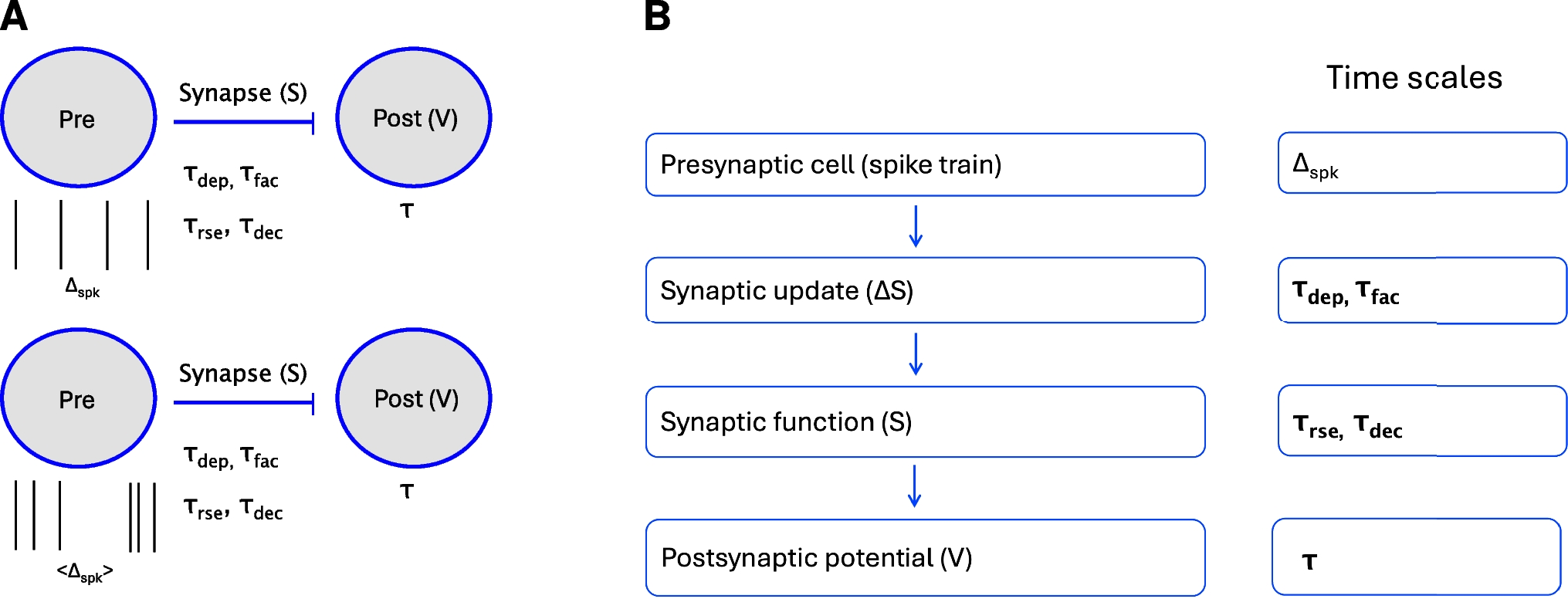
The question we ask in this paper is how the postsynaptic cell’s membrane potential frequency-filters (or -profiles) (curves of the appropriate metrics for each level of organization as a function of the input frequency \(f_\); e.g., Fig. 3-C), depend on the properties of the participating building blocks: the presynaptic spike trains, the synaptic rise and decay dynamics, synaptic short-term plasticity (STP) and the intrinsic properties of the postsynaptic cells (Fig. 1-A).
Specifically, we conduct a systematic study of the steady-state postsynaptic membrane potential (PSP) response to periodic presynaptic inputs over a range of frequencies \(f_ = 1000 / \Delta _\) (Fig. 1-A, top) that capture the PSP filtering properties. We then extend our study to include jittered periodic inputs over a range of mean frequency \(f_ = 1000 / \Delta _\) and Poisson-distributed presynaptic inputs over a range of mean rates \(r_ = 1000 / <\Delta _>\) (Fig. 1-A, bottom).
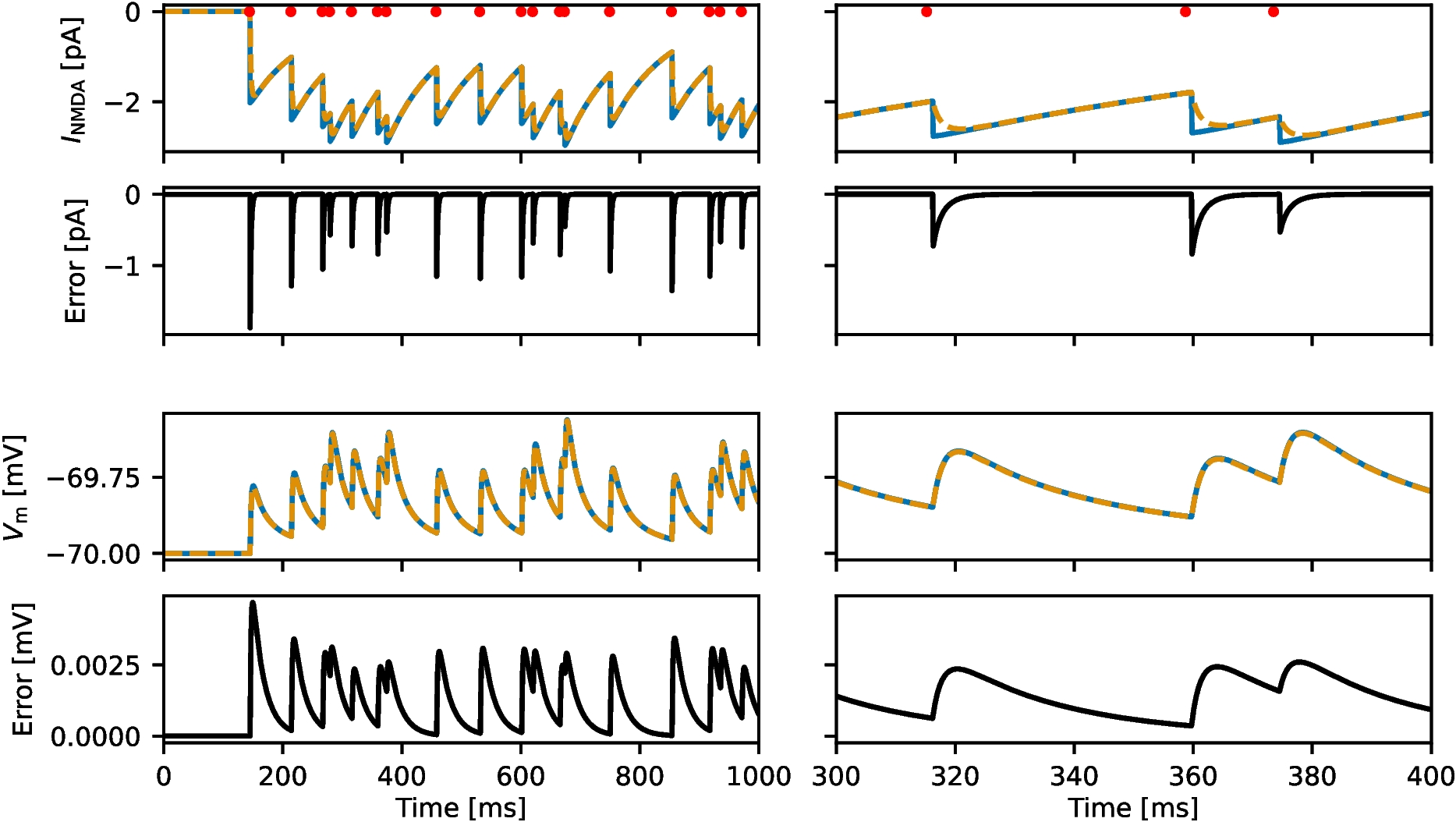
We divide our study in three steps (Fig. 1-B): (i) the response profiles of the synaptic update \(\Delta S\) to the presynaptic spike trains, (ii) the response profiles of the synaptic variable \(S\) to \(\Delta S\), and (iii) the response profiles of the postsynaptic membrane potential \(V\) to \(S\). Synaptic short-term plasticity (STP) operates at the \(\Delta S\) level. The interaction between depression and facilitation (time constants \(\tau _\) and \(\tau _\), respectively) creates the synaptic update sequences \(\Delta S_n = X_n Z_n\) (Fig. 3, left column, blue dots) where \(X_n\) and \(Z_n\) are the sequence of peaks for the depression and facilitation variables \(x\) and \(z\), respectively. These sequences are the target for the synaptic variables \(S\) during the rise phase after the arrival of each presynaptic spike. In the absence of STP, \(\Delta S_n\) is constant (typically set up to one). The interplay of \(\Delta S_n\) and the synaptic dynamics (rise and decay time constants \(\tau _\) and \(\tau _\), respectively) creates the response synaptic (\(S\)) patterns (Fig. 3, middle column). The synaptic variable \(S\) is the input to the current-balance equation (1) where the synaptic patterns interact with the postsynaptic biophysical membrane time constant \(\tau\) to generate the postsynaptic (\(V\)) response patterns (Fig. 3, right column).
Here we focus on the frequency filtering properties of the steady-state responses for \(\Delta S_n\), \(S\) and \(V\). We characterize them by using the \(\bar\), \(\bar\) and \(\bar\) peak profiles (Fig. 3-C, blue), defined as the curves of the stationary peaks for the corresponding quantities as a function of the input frequency \(f_\), and the stationary peak-to-trough amplitude profiles (Fig. 3-C, light blue), consisting of the peak-to-trough amplitude curves as a function of \(f_\), for the latter two quantities.
The temporal filtering properties (transient responses to spike-spike trains) of these feedforward networks were systematically investigated in Mondal et al. (2022).
3.1 \(\bar S\) band-pass filters: interplay of low-pass (depression) and high-pass (facilitation) filtersFrom Eqs. (12), (13) and (5), \(\bar\) is monotonically decreasing (low-pass filter; LPF), transitioning from \(\bar = 1\) (\(f_=0\)) to \(\bar = 0\) ( \(f_ \rightarrow \infty\)), and \(\bar\) is monotonically increasing (high-pass filter; HPF), transitioning from \(\bar = a_f\) (\(f_=0\)) to \(\bar = 1\) (\(f_ \rightarrow \infty\)). This is illustrated in Fig. 4 (red and green) for representative parameter values. The interplay of depression and facilitation produces \(\bar = \bar \bar\) LPFs, BPFs or more complex patterns depending on the relative values of \(\tau _\) and \(\tau _\) (Fig. 4, blue), for fixed realistic values of the remaining parameters.
3.1.1 BPFs: A trade-off between depression- and facilitation-dominated regimesTo simplify the mechanistic analysis, we define
$$\begin \hat_ = \frac}} \hspace \text \hspace \eta _ = \frac}}. \end$$
(43)
Substitution into Eqs. (12) and (13) (with \(x_ = 1\) and \(z_ = 0\)) yields
$$\begin\widehat X=\frac_}\,}_}}&\mathrm&\widehat Z=\frac_\,\eta_}}.\end$$
(44)
This rescaling allows us to investigate the mechanisms of generation of \(\bar\) band-pass filters (BPFs) as a function of a single parameter (\(\eta _\)) describing the relative magnitudes of the single event time constants \(\tau _\) and \(\tau _\). Specifically, \(\hat\) decreases with increasing values of \(\hat_\) in a \(\eta _\)-independent manner, while the rate of increase of \(\hat\) with \(\hat_\) depends on the ratio \(\eta _\) of \(\tau _\) and \(\tau _\). The shapes of the \(\bar\) filters for all values of \(\tau _\) and \(\tau _\) unfold from the shapes of the corresponding \(\hat\) filters by reversing the rescaling.
For large enough values of \(\eta _\) (depression-dominated regime), the increase of \(\hat\) with increasing values of \(f_\) is much slower than the decrease of \(\hat\) (in the limiting case \(\eta \rightarrow \infty\), \(\hat \sim a_f\), a constant). Therefore, \(\hat\) is a LPF (e.g., Fig. 4-A and -B, blue). For small enough values of \(\eta _\) (facilitation dominated regime), \(\hat\) increases very fast with increasing values of \(f_\) as compared to \(\hat\) (in the limiting case \(\eta _ \rightarrow 0\), \(\hat\) increases instantaneously and is approximately a constant, \(\hat \sim 1\)). Therefore, \(\hat\) is also a LPF.
The transition between these two LPFs as \(\eta _\) changes occurs via the development BPFs (e.g., Fig. 4-D to -F, blue). Within some range of values of \(\eta _\) in between the LPFs and BPFs, the \(\bar\) patterns develop a local minimum preceding the local maximum (e.g., Fig. 4-C, blue).
Fig. 3
Representative temporal patterns for the synaptic update \(\Delta S\), the synaptic variable \(S\), and the postsynaptic membrane potential \(V\) in the presence of short-term dynamics (STD). We used the model for the postsynaptic cell described by Eqs. (1)-(4) with STD described by the DA model (7)-(9), and periodic presynaptic spike trains with frequency \(f_\) (see schematic Fig. 1, left). Left column. Short-term dynamics. The peak sequence \(\Delta S_n\) (\(n = 1, \ldots , N_\)) is the synaptic update to the synaptic variable \(S\) upon the arrival of each presynaptic spike, and results from the combined effect of the depression (x) and facilitation (z) variables. The stationary value of the \(\Delta S_n\) sequences is referred to as \(\bar\). Middle column. Synaptic dynamics. The amplitude \(\Gamma _S =\bar_-\bar_\), where \(\bar = \bar_\) and \(\bar_\) are the stationary values of the sequences \(S_\) and \(S_\), respectively. Right column. Membrane potential dynamics. The amplitude \(\Gamma _V = \bar_-\bar_\), where \(\bar = \bar_\) and \(\bar_\) are the stationary values of the sequences \(V_\) and \(V_\), respectively. A. \(f_ = 20 Hz\). B. \(f_ = 40 Hz\). C. Frequency profiles of the stationary peaks \(\bar\) (left, blue), \(\bar\) (middle, blue) and \(\bar\) (right, blue) for the peak sequences \(\Delta S_n\), \(S_n\) and \(V_n\), respectively, and stationary peak-to-trough amplitude profiles \(\Gamma _S\) (middle, light blue) and \(\Gamma _V\) (right, light blue) for \(S\) and \(V\), respectively. The black dots correspond to the presynaptic input frequencies in \(A\) and \(B\). We used the following additional parameter values: \(a_d = 0.1\), \(a_f = 0.1\), \(x_ = 1\), \(z_ = 0\), \(\tau _ = 400\), \(\tau _ = 400\), \(\tau _ = 0.1\), \(\tau _ = 10\), \(C = 1\), \(E_L = -60\), \(G_L = 0.1\), \(I_ = 0\), \(G_ = 0.025\), \(E_ = 0\)
Fig. 4
\(\bar\) filters in response to periodic presynaptic spike inputs (frequency \(f_\)) for the DA model: representative examples. We used Eqs. (12) and (13). A. \(\tau _ = 1\). B. \(\tau _ = 100\). C. \(\tau _ = 200\). D. \(\tau _ = 500\). E. \(\tau _ = 1000\). F. \(\tau _ = 10000\). We used the following additional parameter values: \(a_d = 0.1\), \(a_f = 0.1\), \(x_ = 1\), \(z_ = 0\) and \(\tau _ = 1000\)
3.1.2 From single-event time constants to frequency filters: Control of the filters’ shape by \(}\) and \(}\)The shapes of the \(\bar\) LPFs and \(\bar\) HPFs, and therefore the \(\bar\) BPFs, are controlled by the time constants \(\tau _\) and \(\tau _\) operating at the “single-event” level, which govern the depression and facilitation dynamics, respectively, in response to each presynaptic spike. This is done by the communication of the single event time constants to the global time constants characterizing the temporal filters in response to repeated presynaptic inputs (Mondal et al., 2022) in a presynaptic frequency-dependent manner. The \(\bar\), \(\bar\) and \(\bar\) frequency filters are the steady-states of said temporal filters across presynaptic input frequencies.
We characterize the properties of the \(\bar\) and \(\bar\) filters in terms of the characteristic frequencies \(\sigma _\) and \(\sigma _\), respectively (Fig. 5-A2). These attributes are defined as the frequencies for which the filters reached 63% of the gap between their values at \(f_ = 0\) and \(f_ \rightarrow \infty\) (black dots in Figs. 5-A1 and -A2). For the characterization of the \(\bar\) BPFs we use four attributes (Fig. 5-A3): the characteristic frequencies \(\kappa _\) and \(\kappa _\), the \(\bar\) resonant frequency \(f_,res}\) and the peak frequency \(\bar_\). The characteristic frequencies were computed as the frequency difference between the peak and the frequency value at which \(\bar\) reached 63% of the gap between the peak and the value at \(f_ = 0\) (\(\kappa _\)) and \(f_ \rightarrow \infty\) (\(\kappa _\)). The difference \(\Delta \kappa = \kappa _ - \kappa _\) is a measure of the spread of the BPFs.
Figure 5-B shows the dependence of the characteristic frequencies for the \(\bar\) and \(\bar\) filters \(\tau _\) and \(\tau _\) (for representative values of \(a_d\) and \(a_f\)). Specifically, \(\sigma _\) and \(\sigma _\) are decreasing functions of \(\tau _\) and \(\tau _\), respectively, and decreasing functions of \(a_d\) and \(a_f\), respectively. In other words, the larger the time constants, the more pronounced the decrease and increase of the corresponding filters with \(f_\).
Figure 5-C shows the dependence of the attributes for the \(\bar\) BPFs with \(\tau _\) and \(\tau _\). We fixed the value of \(\tau _ = 1000\) (Fig. 5-C, blue) so the range of resonant frequencies \(f_,res}\) is relatively low. Using this information one can obtain the dependences for other values of \(\tau _\) by reversing the rescaling (43). For comparison, we also present the results for \(\tau _ = 250\) (Fig. 5-C, red). Specifically, the resonant frequency (\(f_\)) decreases with increasing values of \(\tau _\) and \(\tau _\), the \(\bar\) peak increases with increasing values of \(\tau _\) and decreases with increasing values of \(\tau _\) and the peak becomes sharper (\(\Delta \kappa\) decreases) as \(\tau _\) or \(\tau _\) increase. Figure 5-D shows the same results as a function of the characteristic frequency \(\sigma _\).
Fig. 5
\(\bar\) filters in response to periodic presynaptic spike inputs (frequency \(f_\)) for the DA model: frequency attributes. A. The black dots on the \(\bar\), \(\bar\) and \(\bar\) filters indicate the characteristic frequencies (projections on the \(f_\) axis) defined as the change in the corresponding quantities by 63 % of the gap between their final and initial values (\(\sigma _\) and \(\sigma _\)) and between their maximum and minimum values (\(\kappa _\) and \(\kappa _\)). The \(\bar\) resonant frequency \(f_\) is the peak frequency and \(\Delta S_\) is the peak value. B. Dependence of the \(\bar\) and \(\bar\) attributes (characteristic frequencies \(\sigma _\) and \(\sigma _\)) with the depression and facilitation time constants \(\tau _\) and \(\tau _\), respectively. C. Dependence of the \(\bar\) attributes with \(\tau _\) for representative values of \(\tau _\). D. Dependence of the \(\bar\) attributes with \(\sigma _\) for representative values of \(\tau _\). For \(\tau _ = 1000\), \(\sigma _ \sim 17.6\), and for \(\tau _ = 250\), \(\sigma _ \sim 70.1\)
3.2 Interplay of \(}\) and \(}\) filters: inherited and cross-level mechanisms of generation of \(}\) BPFsWe characterize the steady state response profiles of \(S\) to periodic presynaptic inputs by considering two attributes: the steady-state value \(\bar\) (\(=\bar_\)) of the peak sequence \(S_n\) (\(= S_\)) (Fig. 3, middle column, coral dots) and the peak-to-trough steady-state amplitude
$$\begin \Gamma _S =\bar_-\bar_ \end$$
(45)
where \(\bar_\) is the steady-state value of the trough sequence \(S_\) (Fig. 3, middle column, acquamarine dots). Figure 3 (middle column) illustrates that both \(\bar\) and \(\Gamma _S\) vary with the input frequency \(f_\). The temporal filtering properties of \(S\) using these two attributes were investigated in Mondal et al. (2022).
3.2.1 The to-\(}\) synaptic update model with instantaneous \(S\) riseHere we use the approximate to-\(\bar\) model (21) for the dynamics of the synaptic variable \(S\) under the assumption of instantaneous rise time. By construction, the steady state value \(\bar\) is given by
$$\begin \bar = \bar \end$$
(46)
for all input frequencies \(f_\) (\(\bar\) is the steady-state profile of the sequence \(\Delta S_n\)).
The \(}\) filtering properties are inherited from the synaptic update levelThe peak envelope profiles \(\bar\) (\(\bar_\)) are identical to the \(\bar\) profiles for all input frequencies \(f_\). In the absence of STP, \(\bar\) is a constant (\(=1\)), while in the presence of STP, \(\bar\) inherits the filtering properties from the synaptic update level discussed above.
In order to calculate \(\Gamma _S\), one needs to solve the differential equation (21) during the presynaptic ISIs (\(\Delta S_ = \Delta S_\)) and update the solution at the occurrence of each presynaptic spike at \(t = t_\) (\(n = 1, \ldots , N_\)), thus arriving to the following discrete linear difference equation for the trough sequences
$$\begin S_ = e^/\tau _}\, \Delta S_n. \end$$
(47)
Therefore, the peak-to-trough envelope amplitude profile \(\Gamma _S\) is given by
$$\begin \Gamma _S = \bar\, (\, 1 - e^/\tau _}\, ). \end$$
(48)
This expression is the product of two frequency-dependent processes: the \(\bar\) profile and
$$\begin Q_A = 1 - e^/\tau _}, \end$$
(49)
which is a LPF. The shape of the \(\bar\) profile depends on the presence and properties of STP. In the absence of STP, the \(\bar\) profile is independent of \(f_\) and \(\Gamma _S\) is a LPF.
As \(f_\) increases, the \(\Gamma _S\) profiles transition from \(\Gamma _S = \bar\) (\(f_ \rightarrow 0\)) to \(\Gamma _S = 0\) (\(f_ \rightarrow \infty\)) (Fig. 6). For fixed values of \(f_\), the \(\Gamma _S\) profiles transition from \(\Gamma _S = \bar\) (\(\tau _ \rightarrow 0\)) to \(\Gamma _S = 0\) (\(\tau _ \rightarrow \infty\)) as \(\tau _\) increases. In other words, for small enough values of \(\tau _\), the \(\Gamma _S\) profiles reproduce the \(\bar\) profiles (Fig. 6-A1 to A3), but for larger values of \(\tau _\), the \(\Gamma _S\) and \(\bar\) profiles are different. These differences increase as \(f_\) and \(\tau _\) increase (Fig. 6). For generality, in Fig. 6 we included values of \(\tau _\) beyond the biophysically plausible regime for AMPA excitation.
Inherited mechanisms of generation of \(\varvec\) BPFsWhen \(\bar\) is constant (frequency-independent, no STP) or is an LPF, \(\Gamma _S\) is a LPF (Fig. 6-A2 and A3). These LPFs become more pronounced as \(\tau _\) increases. When \(\bar\) is a BPF, the \(\Gamma _S\) BPFs evoked by \(\bar\) become sharper as \(\tau _\) increases (Fig. 6-A1). These \(\Gamma _S\) filters are inherited from the \(\bar\) ones and modulated by \(\tau _\).
Cross-level mechanisms of generation of \(\varvec\) BPFsIn contrast, when \(\bar\) is a HPF, \(\Gamma _S\) BPFs emerge as the product of a HPF and a LPF (Fig. 6-A4). As \(\tau _\) increases, the \(Q_A\) LPF is more pronounced as a function of \(f_\) and therefore the \(\Gamma _S\) BPF is sharper and peaks at a smaller value (compare Fig. 6-A4, -A5 and -A6).
3.2.2 The to-\(}\) synaptic update model with non instantaneous \(S\) riseHere we focus on the effects of the synaptic rise time on the generation and modulation of synaptic filters, particularly synaptic BPFs. We use the approximate model (22) for the dynamics of the synaptic variable \(S\) when the assumption of instantaneous rise is relaxed. The solution to the first and second terms in (22) are given by
$$\begin S = \Delta S + [\, S(t_) - \Delta S \,]\, e^)/\tau _} \end$$
(50)
and
$$\begin S = [\, \Delta S + (\, S(t_)-\Delta S\, )\, e^/\tau _})\, ]\, e^-T_)/\tau _}, \end$$
(51)
respectively. Using this, one can compute the difference equation governing the evolution of the sequence of peaks
$$\begin S_ = \Delta S\, \left( \, 1 - e^/\tau _}\, \right) + S_n\, e^-T_)/\tau _}\, e^/\tau _}. \end$$
(52)
By assuming a constant \(\Delta _ = \Delta _\), one obtains
$$\begin \bar = \bar\, \frac/\tau _}}/\tau _}\, e^-T_)/\tau _}\, } \end$$
(53)
and
$$\begin \Gamma _S = \bar\, \left[ 1 -e^-T_)/\tau _} \right] . \end$$
(54)
Both expressions are the product of frequency-dependent filters and reduce to Eqs. (46) and (48) for \(\tau _ \rightarrow 0\) and \(T_ \rightarrow 0\) with \(\tau _ / T_ \ll 1\).
We first focus on the \(\bar\) profiles. For \(\tau _ \rightarrow 0\), the second factor in Eq. (53)
$$\begin Q_C = \frac/\tau _}}/\tau _}\, e^-T_)/\tau _}\, } \end$$
(55)
approaches \(Q_C = 1\) for all \(f_\) and therefore the \(\bar\) profiles are approximately equal to the \(\bar\) profiles. For \(\tau _> 0\), \(Q_C\) is a HPF and therefore the \(\bar\) profiles depart from the \(\bar\) profiles
Specifically, for \(\tau _> 0\), \(Q_C\) changes from \(Q_ = 1 - e^/\tau _} < 1\) (for \(f_ = 0\)) to\(\beginQ_=(1-e^/\tau_})/\end\) \((1-e^(1/\tau_-1/\tau_)})>1\) (as \(f_ \rightarrow \infty\)). \(Q_C = 1\) for \(f_ = 1000/T_\), independently of \(\tau _\) and \(\tau _\). As \(\tau _\) increases (all other parameters fixed), within some bounds, \(Q_\) decreases and \(Q_\) increases, causing an increase in the HPF amplitude of \(Q_C\). As \(\tau _\) increases (all other parameters fixed), also within some bounds, \(Q_C\) increases for \(0< f_ < 1000/T_\) and decreases for \(f_> 1000/T_\).
Attenuation of the \(\varvec}\) filters (BPFs, LPFs and HPFs) inherited from the synaptic update levelTherefore, for \(f_ < 1000/T_\), increasing values of \(\tau _\) cause an attenuation of the \(\bar\) profiles (Fig. 7-A1 to -A3), and this attenuation is less pronounced the larger \(\tau _\) (not shown). For \(f_> 1000/T_\), increasing values of \(\tau _\) cause an amplification of the \(\bar\) profiles (Fig. 7-A3), which is less pronounced the larger \(\tau _\) (not shown). However, for \(T_ = 1\), the latter range is well beyond the frequencies we are interested in this paper.
The bounds mentioned above are set by the requirement that the denominator of \(Q_C\) is positive, which in turn requires that \(\Delta _> T_ (\tau _-\tau _)/\tau _\). This is satisfied for all values of \(\Delta _\) if \(\tau _ < \tau _\). (For larger values of \(\tau _\), this imposes a bound on \(\Delta _\) for which \(Q_C> 0\).) The realistic values of \(\tau _\) and \(\tau _\) we use here satisfy this condition. Moreover, for these values of \(\tau _\) and \(\tau _\), \(Q_C\) is a HPF, converging asymptotically to \(Q_\).
Cross-level mechanisms of generation of \(\varvec}\) BPFs and attenuation of \(\varvec}\) filtersBecause \(Q_C\) has HPF properties, the question arises whether a \(\bar\) BPF can be created by the interplay of a \(\bar\) LPF and \(Q_C\) for nonzero values of \(\tau _\). (For instantaneous rise times, \(\bar\) BPFs can only be inherited since \(\bar = \bar\); see Section 3.2.1). Figures 7-A4 to -A7 illustrates that this is indeed possible \(\tau _> 0\). The generation of a \(\bar\) band-pass filter requires that the \(Q_C(0)\) is low enough, which is achieved by increasing \(\tau _\) above some threshold value (Fig. 7-A5). This BPF can be amplified by making the increase of \(Q_C\) sharper for low values of \(f_\), which can be achieved by increasing \(\tau _\) (Fig. 7-A6). The BPF in Fig. 7-A5 is attenuated by further increasing \(\tau _\) since this causes the intersection between the constituents low- and high-pass filters to move down (Fig. 7-A7). An increase in the values of \(\tau _\) (Fig. 7-A7) causes the \(\bar\) LPF to decrease sharper as compared to Fig. 7-A5, decreasing the intersection between the \(\bar\) LPF and the \(Q_C\) HPF. The resulting attenuation produces a \(\bar\) LPF (Fig. 7-A7).
Cross-level mechanisms of generation of \(\varvec\) BFPs and attenuation of \(\varvec\) filtersThe second factor in Eq. (54),
$$\begin Q_D = 1 -e^-T_)/\tau _}, \end$$
(56)
is a LPF, provided \(\Delta _\) is large enough as compared to \(T_\), and is independent of \(\tau _\). Therefore, the effect of \(\tau _\) on the \(\Gamma _S\) filters is inherited from the effect of \(\tau _\) on \(\bar\) filters. The \(\Gamma _S\) filters are further attenuated by the \(Q_D\) filter. The \(\Gamma _S\) BPFs, in addition, become wider and the \(\Gamma _S\) resonant frequency is displaced (Fig. S14-A1, -A3). The attenuation is more pronounced for the larger frequencies as \(\tau _\) increases, therefore the BPFs become sharper and the \(\Gamma _S\) resonant frequency is displaced as \(\tau _\) increases (not shown).
3.3 Interplay of \(}\) and summation PSP filtersThe synaptic variable \(S\) determined by Eq. (3) is the input to the current balance equation (1) for the postsynaptic voltage response \(V\). In the absence of STP (\(\Delta S = 1\)), summation effects give rise to postsynaptic (PSP) HPFs whose properties depend on the membrane potential properties, particularly the membrane time constant (\(\tau\)). In the presence of STP, the PSP filters reflect the interaction of the \(\bar\) filters and the summation filters. For small enough values of \(\tau\), the PSP filters are well approximated by the \(\bar\) filters. Previous work by other authors has considered PSP and \(\bar\) filters to be proportional (e.g., Markram et al., 1998; Markram et al., 1998b; Markram et al., 1998a, but see Drover et al., 2007). However, for larger values of \(\tau\), the PSP filters are expected to depart from the weak modulation of the \(\bar\) filters.
Here we us the “by-\(\Delta S\)” model (23) described in Section 2.2.2 as an intermediate step for the investigation of the mechanisms that govern the interaction between the \(\bar\) and PSP summation filters. We investigate the response of conductance-based models to periodic presynaptic inputs in the presence of STP in the next Section. The simplified model we study here has the advantage of being amenable to analytical calculations that provide a better insight into the mechanistic aspects of the interaction between the \(\bar\) and PSP summation filters as compared to conductance-based models, and therefore pave the way for the investigation of said models.
3.3.1 The by-\(}\) update model with instantaneous \(S\) riseAs a first step, we focus on instantaneous rise for \(S\). In contrast to the to-\(\bar\) model, instantaneous rise is not a natural assumption when \(S\) is interpreted as the PSP because of the effects of the membrane time constant.
By solving the differential equation (23) for a constant value of \(\Delta S_ = \Delta S_\) during the presynaptic ISIs and updating the solution at each occurrence of the presynaptic spikes at \(t = t_n\), \(n = 1, \ldots\), one arrives to the following discrete linear differential equation for the peak sequences in terms of the model parameters
$$\begin S_ = e^/\tau _}\, S_n+ \Delta S_n. \end$$
(57)
The \(\bar\) profile is given by the steady state values of (57)
$$\begin \bar = \frac S}/\tau _}}. \end$$
(58)
By construction, the \(\Gamma _S\) profiles are
$$\begin \Gamma _S = \bar. \end$$
(59)
In other words, the \(\Gamma _S\) filtering properties are inherited from the \(\bar\) profiles.
The \(\varvec}\) and \(\varvec}\) profiles depart from each other for relatively large values of \(\varvec}\)Equation (58) is the product of two frequency-dependent processes. The factor
$$\begin Q_B = \frac/\tau _}} \end$$
(60)
is a HPF. It transitions from \(Q_B = 1\) (for \(f_=0\)) to \(Q_B \rightarrow \infty\) (for \(f_ \rightarrow \infty\)), and it increases faster the larger \(\tau _\). As already discussed, the shape of the \(\bar\) profile depends on the presence and properties of STP.
From Eq. (58), for small enough values of \(f_\), \(\bar \sim \bar\). In the limit \(f_ \rightarrow 0\), \(\bar = \bar\). As \(f_\) increases, \(Q_B\) increases and therefore the difference between \(\bar\) and \(\bar\) also increases.
\(\varvec}\) BPFs and LPFs transition to \(\varvec}\) HPFs as \(\varvec}\) increases in the presence of STPIn the absence of STP, \(\bar\) is constant and therefore \(\bar\) increases unboundedly as \(f_ \rightarrow \infty\) (Fig. 6-B3). Similarly unbounded \(\bar\) profiles are also obtained for \(\bar\) HPFs. Under certain circumstances, the presence of STP puts a bound on the increase of \(\bar\), particularly for large values of \(\tau _\) and the resulting filters remain bounded. Specifically, \(\bar\) BPFs (Fig. 6-B1) and LPFs (Fig.
Comments (0)