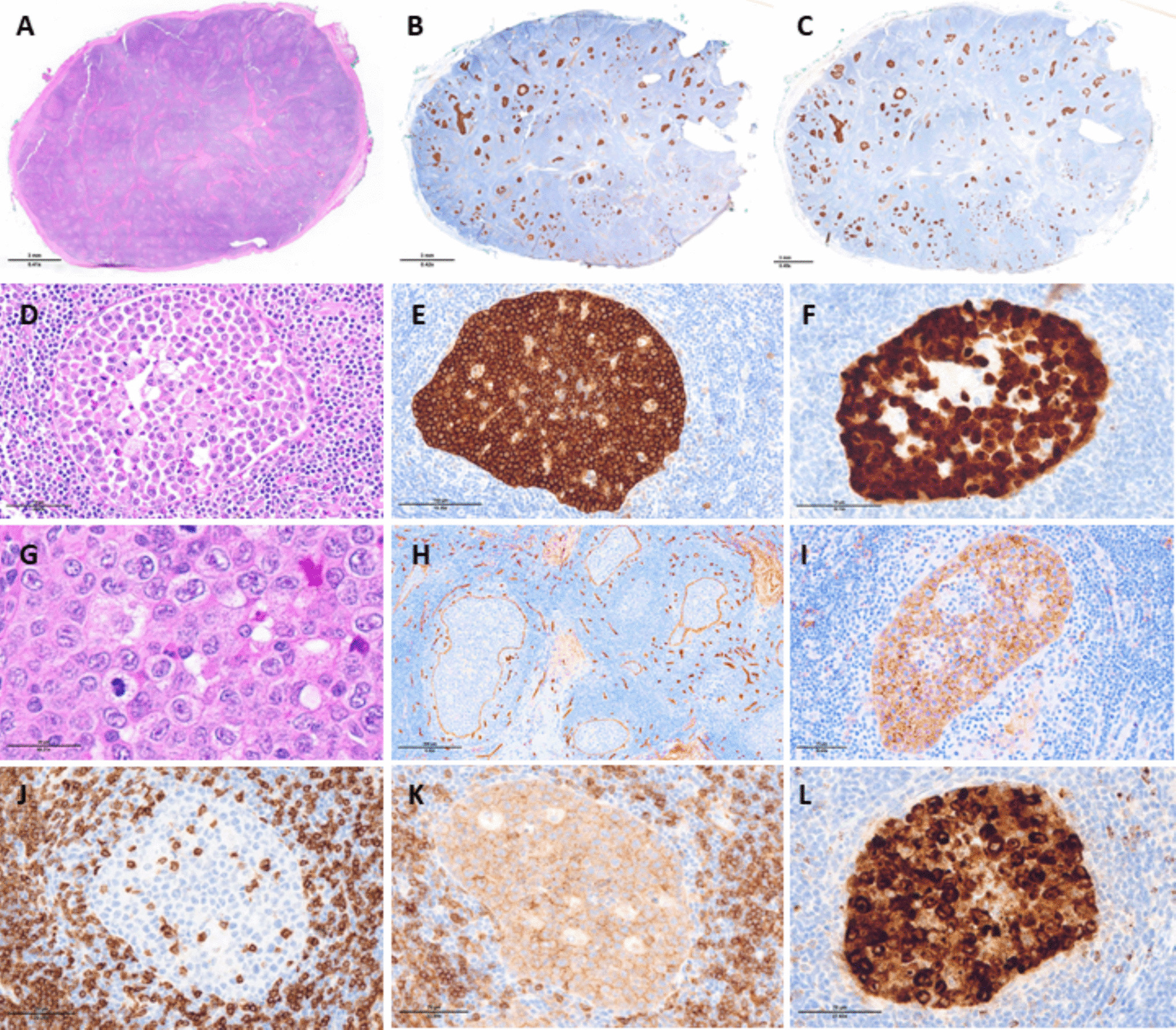
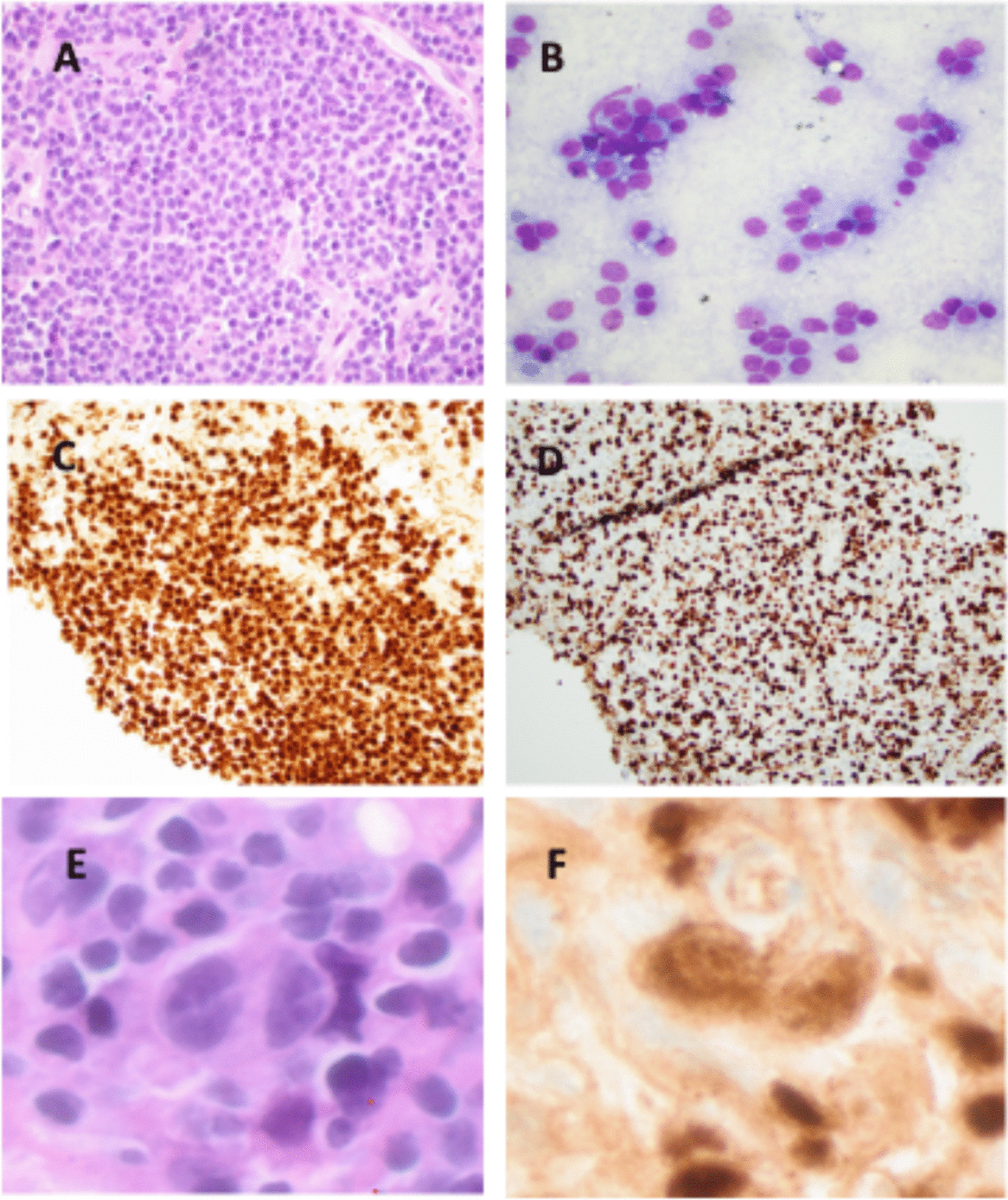
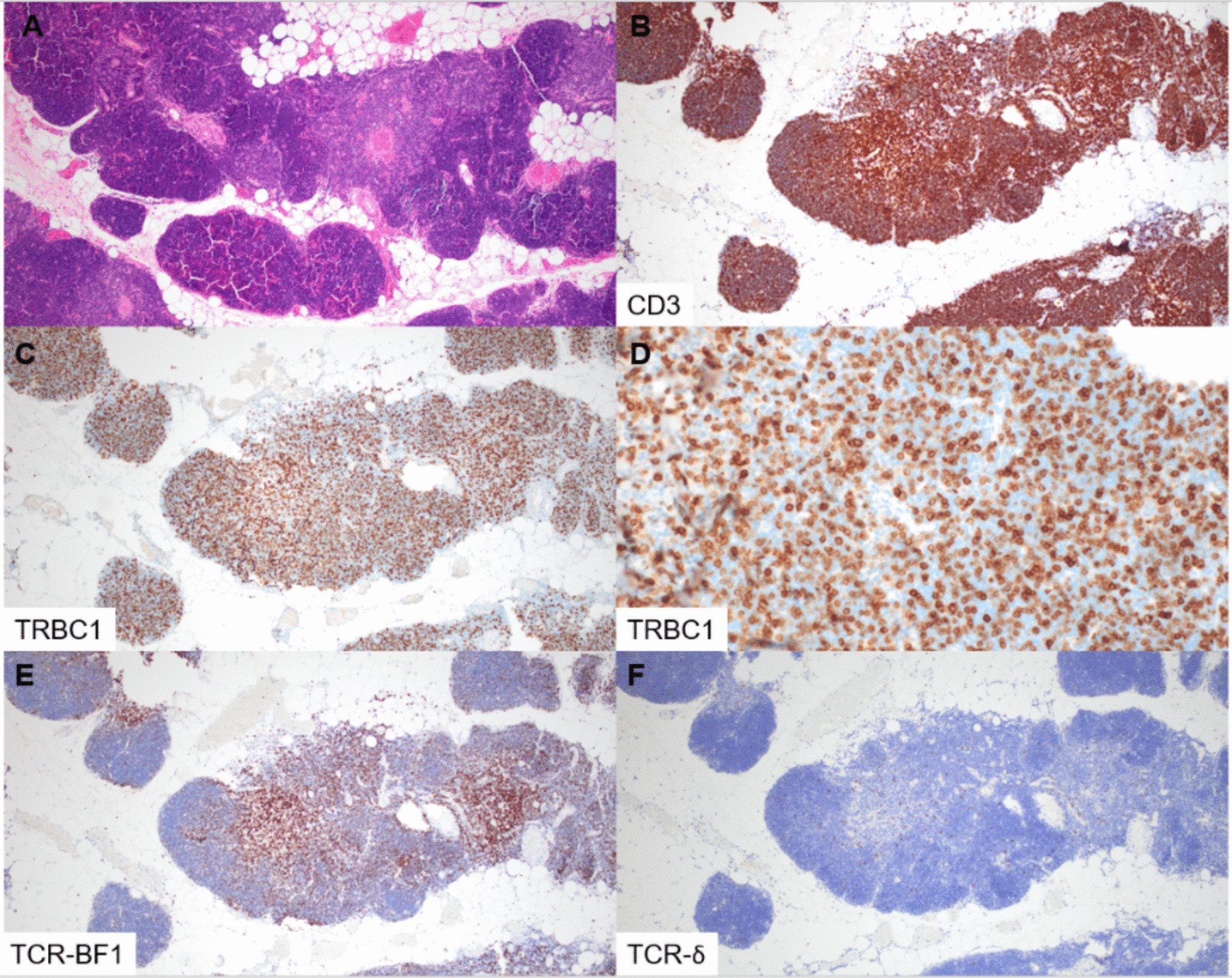
In this study, we examined the expression of TRBC1 by IHC in immature T-cell proliferations. All cases of T-LL showed abnormal, “restricted” TRBC1 staining profiles, whereas benign thymic tissue and thymomas demonstrated a polytypic pattern of TRBC1 expression. Patients with multiple specimens showed consistent TRBC1 staining across tissue sites and sampling time points. TRBC1 IHC also showed excellent staining quality in decalcified bone marrow core biopsies. During the course of our study, a complementary IHC analysis of a similar cohort of immature T-cell populations was published by Lee et al. [17], describing non-restricted TRBC1 patterns in thymic tissue and restricted TRBC1 patterns in T-LLs (19% TRBC + and 81% TRBC1−). Our study corroborates their findings, identifying similar proportions of TRBC1 + (14%) and TRBC1− (86%) cases. As a novel feature, we also describe the phenotypic patterns of TRBC1 staining in relationship to TCR expression, with a large subset of cases showing TCR-δ + or TCR null phenotypes. Molecular methods for clonality assessment (TCR γ PCR, ClonoSEQ) were supportive of a clonal T-cell population in 13 of 15 (87%) tested cases, with the outliers being two cases lacking dominant clonal sequences by ClonoSEQ analysis. One case was an ETP-LL, which is known to have low rates of functional TCR gene rearrangement [18, 19]. The second discrepant case failed DNA quality controls at our institution, raising consideration for specimen quality limitations that may have impacted analysis.
Our study utilized both CD3 and TdT as comparator stains for TRBC1 evaluation in T-LL cases. While CD3 and TdT staining were positive in all cases, the intensity and degree of staining were not uniform, as expected based on the known phenotypic variability that can be seen in T-LL [20]. Accordingly, we found it helpful to have both immunophenotypic markers to delineate the extent and distribution of T-lymphoblast populations. Nevertheless, the TRBC1/CD3 and TRBC1/TdT scoring strategies each posed challenges. In addition to highlighting the immature populations of interest, CD3 stains background mature T cells, confounding evaluation in cases with frequent admixed non-neoplastic lymphocytes. Meanwhile, TdT was more specific for immature lymphoid populations, but tended to be more sensitive to fixation and decalcification effects in our laboratory. We selected cutoffs of ≤ 15% and ≥ 85% TRBC1 staining to define negatively and positively restricted cases, respectively. These thresholds have been repeatedly cited in flow cytometry literature [7,8,9,10,11] and have been employed to good effect in our daily flow cytometry practice. While practical, these cutoffs are admittedly arbitrary. The range of expected TRBC1 positivity shows mild variation across studies, based on the T-cell subsets and sample types analyzed [6, 8]. In our experience with T-LL cases, when the blast population is prominent and associated with tissue effacement, a formal quantitative assessment may not be necessary, as such blast populations are clearly uniformly positive or negative. Conversely, irrespective of scoring strategy, we anticipate that accurate TRBC1 IHC evaluation will be challenging (and in some instances impossible) in samples with low-level involvement by T-LL, particularly bone marrow. In this context, the blast population of interest will be admixed with frequent non-neoplastic cells, and comparator stains (CD3, TdT) are not entirely specific for the blast population of interest. This limitation was previously noted by Zhou et al. in their analysis of TRBC1 staining in mature T-cell lymphomas, in which TRBC1 IHC showed decreased sensitivity in the setting of low tumor burden [13]. In this regard, flow cytometry maintains an advantage over IHC methods, in its ability to detect small aberrant populations by evaluating TRBC expression in conjunction with additional immunophenotypic markers. To develop novel TRBC1% cutoffs for use in FFPE tissue, several groups have explored manual microscopy and digital analysis methods, with each reporting different optimal thresholds for defining positivity and negativity. Lower thresholds for TRBC1 positivity ranged from ≤ 25 to ≤ 36.3% while upper thresholds ranged from ≥ 70 to ≥ 79% [12,13,14, 17]. As our series is exclusively comprised of T-LL cases with high tumor burden and only contains three patients with positively restricted TRBC1, we feel it is not an ideal cohort to establish/validate diagnostic cutoff points.
As the frequencies of TCR and TRBC1 expression observed in our T-LL cohort differ substantially from those of mature T-cell lymphoproliferative disorders, further study of immature T-cell proliferations is warranted. Immunohistochemical assessment for TRBC2 would be an enticing next step, but unfortunately, a TRBC2 antibody suitable for IHC was not commercially available at the time of this study. Accordingly, for the 20 of 24 (83%) cases lacking TRBC1 expression, we could not determine the proportion of TRBC2-positive and TRBC null cases. We hypothesize that the TCR-BF1 +, TRBC1− subset of T-LL cases is the most likely subset to exhibit TRBC2 restriction. However, this should be further interrogated, particularly due to the challenges associated with TCR-BF1 IHC interpretation, which frequently required three-pathologist review (5/24 specimens, 21%) to distinguish dim/subset versus negative staining. This difficulty may stem from the innate biological variability of TCR expression in T-LL, in which expression can be dim, inconsistent across membranous and cytoplasmic cellular compartments, and/or limited to a subset of blasts [11, 21, 22]. In addition, we observed a large proportion of T-LL cases with apparent expression of TCR-δ (29%), and all TCR-δ + cases lacked TRBC1 expression. The significance of this finding is uncertain, as there is only limited data examining TCR-δ expression by IHC in T-LL cohorts. Normal maturing thymocytes show transient TCR-δ expression prior to maturation and adoption of a permanent TCR α/β phenotype [23, 24]. Furthermore, TCR-δ expression has been described in a subset of T-LL cases that lack both surface TCR γ/δ expression and mature γ/δ transcripts [25]. Thus, the mere expression of TCR-δ may not entirely correlate with a surface TCR γ/δ + phenotype, which is reported in only a small proportion of T-LLs (~ 9–12%) by flow cytometry [21, 26,27,28,29]. Indeed, for the one TCR-δ + case for which flow cytometry correlation could be performed (patient 11), surface TCR γ/δ expression was not observed.
The ability to correlate IHC staining with flow cytometric immunophenotyping was limited in this retrospective study, as the specific panels of evaluated flow cytometry markers varied across specimens. Surface TRBC1, TCR γ/δ, and/or TCR α/β assessments were included in only a minor subset of flow cytometry analyses, and cytoplasmic evaluation of these markers was not performed. Existing flow cytometry literature is also limited in this regard. The TRBC phenotype of T-LL has only been described for small case cohorts, with studies primarily reporting the profile of TRBC1, without concomitant assessment of TRBC2 [11, 15, 30, 31]. A paired analysis comparing the assessment of TRBC1, TRBC2, TCR α/β, and TCR γ/δ by immunohistochemical and flow cytometry methods (to include evaluation of surface and cytoplasmic expression) would be invaluable for establishing the spectrum of TCR/TRBC profiles in T-LL. TCR and TRBC transcriptional profiling is an additional technique that could be explored, but is beyond the scope of the current investigation.
While TRBC1 IHC is emerging as a useful tool for establishing clonality, a variety of morphologic, immunophenotypic, and molecular-cytogenetic features can be utilized to identify aberrant T-lymphoblast populations, and the primary diagnosis of T-LL is often straightforward when tissue sampling is adequate. However, TRBC1 IHC could be of added diagnostic value in select situations. For example, when evaluating specimens from the mediastinum, distinguishing between benign thymic hyperplasia, thymoma, and T-LL can be challenging, particularly in the setting of limited tissue and/or the absence of conclusive flow cytometric immunophenotyping. A limited IHC panel including CD3, TdT, TRBC1, and keratin may be sufficient to distinguish these entities, with the utilization of minimal tissue. Such a strategy would also be helpful for accurately differentiating ectopic thymic tissue from T-LL when evaluating small samples from the head and neck region. TRBC1 IHC could have utility in distinguishing indolent T-lymphoblastic proliferations from T-LL, as long as the lymphoblast population of interest is sufficiently prominent to be visualized/localized on serial tissue sections. Finally, the selective utilization of TRBC isoforms by T-cell neoplasms presents a compelling potential therapeutic target. An intervention that targets one TRBC isoform has selective activity against the neoplastic T-cell population of interest, while preserving sufficient background T cells to maintain host immunity [32,33,34]. In this setting, TRBC phenotyping by IHC may have utility in appropriately selecting patients for TRBC-targeted therapy.
In summary, TRBC1 expression assessment by IHC represents a viable alternative to flow cytometry for the evaluation of immature T-cell populations. T-LL cases with high tumor burden showed overtly restricted patterns of TRBC1 expression, whereas benign thymic tissue and thymomas consistently showed polytypic staining patterns. TRBC1 IHC may be particularly helpful in distinguishing between benign thymic tissue and T-LL when evaluating small samples from the mediastinum and head and neck. However, diagnostic utility is anticipated to be more limited in situations of low tumor burden. In this small series, TRBC1 IHC and molecular methods of clonality showed high concordance (87%), although TRBC1 was slightly more sensitive in identifying clonal populations. Our study demonstrates that patterns of TCR and TRBC1 expression are notably different in malignant T-lymphoblast populations compared to mature T-cell lymphomas, while highlighting the need to further refine the spectrum of TCR and TRBC expression in immature T-cell proliferations, with particular emphasis on exploring TRBC2 + and TRBC null phenotypes.

Comments (0)