
Remember me

The micromeres in Tubifex embryos lineage-specifically contribute to the head epidermis, cerebral ganglion, buccal cavity, pharynx, oesophagus, and yolk sac ectoderm of the progeny, comprising the dorsal temporary epithelium and ventral/ventrolateral epidermis (Table 1). No evidence currently shows that the cells derived from either ectodermal or mesodermal teloblasts are distributed in these structures [28, 38, 42]. Thus, these structures (tissues and organs) are each exclusively produced by the descendants of specific micromeres.
We found that each micromere contributed the progeny to either the left or right side of a specific structure (Table 1). Each Tubifex organ is likely composed of the descendants of two or more different micromeres. The cerebral ganglion consists of cells derived from 1c (right) and 1d (left). However, we failed to detect labelled cells in the injected embryos on the right side of the buccal cavity, right side of the pharynx, or left side of the oesophagus (Fig. 6c, f, h, m). This indicated that the cells located in these portions remained unlabelled in all injected embryos. These unlabelled cells could have derived from the B-quadrant micromeres (2b and 3b) that were not injected with the lineage tracer in this study.
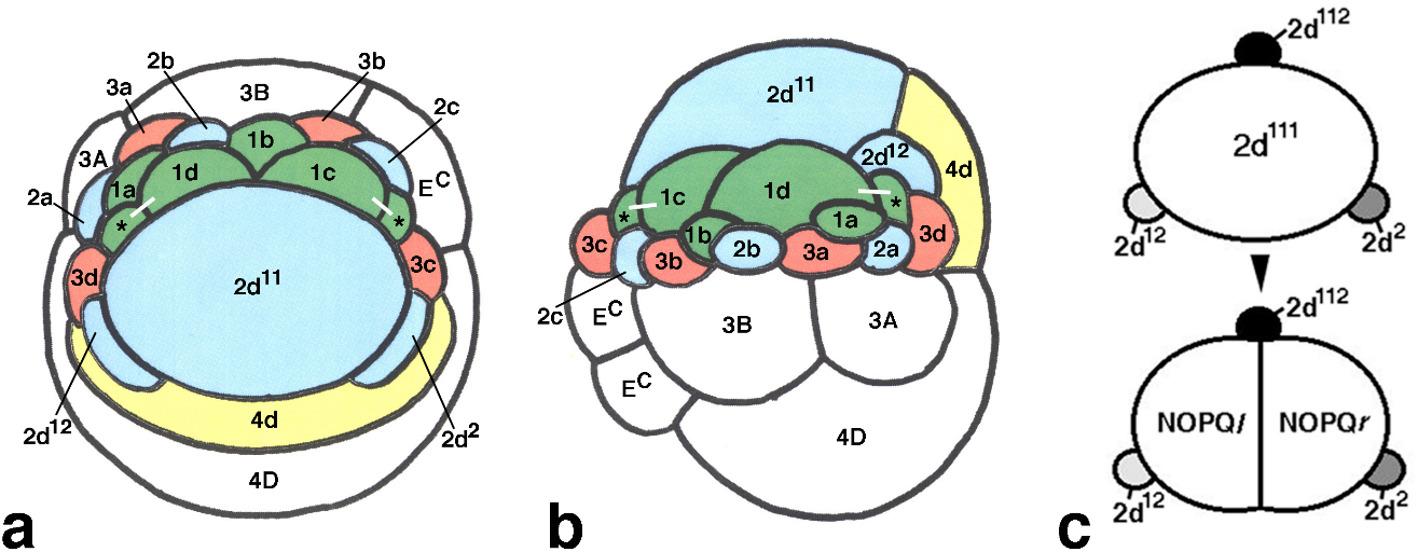
Table 1 Summary of micromere fate mapping in T. tubifexThe smaller micromeres in Tubifex embryos are bilaterally symmetrically arranged during early development. The micromeres located to the left of the midline at the 22-cell stage generate the left side of the embryo in terms of tissues and organs, and those located to the right of the midline give rise to the right side of the embryo based on the position of individual micromeres relative to the (future) midline (Fig. 1a). The former micromeres corresponding to the left side of the embryo include 1a, 1d, 2a, 3a, 3d, and 2d12, which are referred to as leftward micromeres, and the latter micromeres corresponding to the right side consist of 1b, 1c, 2c, 3c, and 2d2, which are referred to as rightward micromeres (Table 1). This pattern of micromere contribution to the embryo is similar to that in other spiralians, including polychaete annelids Platynereis dumerilii and Capitella teleta, gastropod molluscs Crepidula fornicata, Ilyanassa obsoleta and Patella vulgata, and the nemertean Cerebratulus lacteus, in which micromeres organise in a dome-shaped assemblage. Micromeres 1a, 1d, 2a, 3a, and 3d correspond to the left side, and micromeres 1b, 1c, 2c, 3b, and 3c correspond to the right side of the larval body plan in these spiralians, except for 1b and 1d in Ilyanassa, which correspond to the ventral and dorsal sides, respectively, rather than the left/right axis (see [43, 44] for reviews). These spiralians are distinct from Tubifex in that individual micromeres are radially symmetrically arranged, even when the fourth quartet micromeres form [44]. This radial symmetry is disrupted only when specific blastomeres such as 2d, 3D, and 4d function as embryonic organisers in Capitella, Ilyanassa, Lymnaea, and Crepidula; if such organiser activities are abrogated, the resulting larvae fail to show any signs of bilaterality and remain radialised [45–51].
The bilateral symmetry in Tubifex with respect to the future midline is established in the micromere arrangement at the time of 4d formation (Fig. 1a). The large 2d micromere, which forms at the animal pole of the embryo, plays a central role in generating this bilateral symmetry. However, this micromere likely does not emanate a signal to induce bilaterality in the micromere assemblage, as in other spiralians. The 2d micromere rather serves as a mould that supports the arc shape of the micromere assemblage. However, micromere fates may be specified through some inductive signals emanating from micromere 2d and/or its derivatives, as in the case of Capitella [49], because the developmental fates of all micromeres are likely not autonomously determined by inherited factors. Eye specification in Ilyanassa embryos involves inherited factors and positions with respect to the inducing D macromeres [51]. Micromere 2d11 (the larger grand-daughter of 2d) in Tubifex functions together with 4d as an annelid axial organiser [52]. However, little is known about the impact of these organisers on the micromeres.
4.1 Comparison with previous findings on Tubifex embryogenesisThe cerebral ganglion in Tubifex is thought to derive from the ectodermal GB [19]. This view is mainly based on Penners’s description of the anterior-most part of the GB including the rudiment of the cerebral ganglion (Penners 1924, p. 276) [53]. However, no data support this view. We found that the rudiment of the cerebral ganglion emerges within the anteriorly located micromere association (Fig. 2a) but not in the GB. We found that the cerebral ganglion in Tubifex was composed of descendants of two micromeres, 1c and 1d. Furthermore, no labelled progeny cells were found in the regions corresponding to the cerebral ganglion when lineage tracers, such as DiI and HRP, were injected into NOPQ or 2d111 (see Fig. 1c) [28, 29, 42]. These findings indicate that the cerebral ganglion does not develop from the ectodermal GB.
The fate of micromeres that produce the yolk sac ectoderm is not thoroughly understood. The surface of Tubifex embryos undergoing GB elongation during stages 13–14 is covered with epithelial cell layers, which is the collective yolk sac ectoderm, except for the regions corresponding to the GBs and teloblasts [53, 54]. The yolk sac ectoderm was thought to comprise the progeny of micromeres that persist through embryogenesis to differentiate into a definitive epidermis [15, 53, 54]. We found that the dorsal yolk sac ectoderm was derived from 1c, 1d, and 2d112 and the ventral yolk sac ectoderm from 3c, 3d, 2d2, and 2d12. Descendant cells of these micromeres (other than 1c and 1d) produced a definitive epidermis in the ventral and ventrolateral regions of the embryo. In contrast, the 1c and 1d descendants were undetectable before embryogenesis completion, except for those localised to the dorsal midline (Fig. 5c). The juvenile dorsal and lateral regions are covered with an epidermis derived from ectodermal GBs [42]. These observations suggest that the epithelial cells derived from 1c and 1d in Tubifex are replaced by epidermal cells derived from ectodermal GBs before the end of embryogenesis.
Little is known about the process of the epithelial cells replacement by the GB-derived epidermal cells in Tubifex. Our previous cell lineage studies showed that the dorsalward expansion of the ectodermal and mesodermal layers derived from the GBs was initiated at the anterior-most part of the embryo and progressed in anterior to posterior (Fig. S3c, d) [28]. The contour of the dorsal margin of the expanding mesodermal layer, which is the coelomic wall, was oblique but not parallel to the ventral midline in the anterior half of the embryo in late stage 16 (Fig. S3c, d). The present study showed that the contour of the ventral side of the EL was oblique but not parallel to the ventral midline in the anterior region of the embryo in the late part of stage 16 (Fig. 5b; Fig. S3a, b). A comparison of Fig. S3b with Fig. S3d shows that the EL area was similar to the area that remained uncovered by the mesodermal layer. This finding suggests that the dorsalward retraction of the EL and the dorsalward expansion of the GBs follow a similar time course, at least in the anterior half of the embryo. Based on these considerations, it is tempting to postulate that the GB-derived mesodermal cells affect the micromere-derived epithelial cells and give rise to the ‘disappearance’ of intact epithelial cells. In this regard, it is intriguing to note that the dorsal edges of the coelomic walls migrate between the yolky endoderm and a thin layer of squamous epithelium during the dorsalward expansion of the GBs [28]. At present, it is equally possible that the decrease in the EL area occurs independently of the GBs. Further studies are required to differentiate these possibilities. It would be significant to examine the behaviour of the micromere-derived epithelial cells in embryos where the GBs are prevented from expanding dorsally.
The dorsal and ventral yolk sac ectoderms were also distinct from each other in their positions relative to the GB. The dorsal ectoderm was maintained on the dorsal side of the GB on either side of the embryo during gastrulation (Figs. 3a and b and 8i and j). In contrast, the cell bands comprising the ventral yolk sac ectoderm changed positions relative to the GB. The ventral cell bands were initially between the ventral midline and dorsally located GB on either side of the embryo (Fig. 9a). The cell bands facing the ventral midline, which were those derived from 3c and 3d, exactly superimposed on the GBs (Fig. 8a, c) after gastrulation completed (Fig. 9c), whereas the other cell band ran along the dorsal GB margin on both sides of the embryo (Fig. 8e, g).
Figure 9 shows that the GBs changed positions relative to the ventral midline during rearrangement. However, none of the ventral cell bands derived from 3c, 3d, 2d2, or 2d12 exhibited changes in position relative to the ventral midline. These observations indicate that the ventral cell bands undergo dorsalward movement on the GB, which moves to the ventral midline. The ventral cell bands must move at the same rate as the GBs as the ventral cell bands remained in their initial positions (Fig. 9). In summary, the epithelial cells derived from 3c, 3d, 2d2, and 2d12 may become transiently motile when encountering the pressure of the GB. The ventral surface of the Tubifex embryo in stage 15 exhibits a bilateral pair of arrays of cells that assume a round shape in the anterior half when viewed using scanning electron microscopy (SEM) (Fig. S4) [16]. SEM observations also showed that cells become round in shape around the site where the dorsal margin of the cell band intersects with the ventral margin of the GB (as indicated by the double arrowheads in Fig. S4; also see Fig. 7c). Cells comprising the ventral cell bands may become active or motile around the time these cell bands appear on the GBs. Further studies are required to verify whether the ventral cell band moves along the GB. The behaviour of individual cells in the ventral cell bands can be examined using time-lapse live-cell imaging in combination with live fluorescently labelled GBs.
4.2 Comparison with other annelidsThe Helobdella, Capitella, and Platynereis micromeres contribute progeny to two or more structures such as cells, tissues, and organs (Table 2). Micromeres 2a (a’’) and 2c (c’’) in Helobdella are exceptions, which are fated to die [55]. One or two structures for each micromere are shared by all three annelid species. For example, the 1a progeny contribute to both the head epidermis and brain (supraesophageal ganglion) in Helobdella, Capitella, and Platynereis, whereas the trunk epithelium in Helobdella, the eye in Capitella, and primary trochoblasts in Platynereis are all species-specific (Table 2).
Table 2 Comparison of micromere cell fate data for four annelid speciesTubifex is unique compared with these species in that each micromere appears to contribute progeny to a single structure, except for 1c, 1d, and 2c, which each produce two structures (Table 1). The micromere-derived structures in Tubifex are shared with other annelids, except for 3c and 3d, which contribute progeny to the epidermis on the trunk region. Micromeres 3c and 3d does not contribute to the epidermis in any of the three annelids (Helobdella, Capitella, and Platynereis).
Seven of the twelve Tubifex micromeres examined in this study (1c, 1d, 3c, 3d, 2d2, 2d12 and 2d112) contributed progeny to the trunk region (Table 1). Almost the same set of micromeres in Helobdella contribute their progeny in the trunk region (Fig. 12) [55]. Nine micromeres (1c, 1d, 2a, 2c, 3c, 3d, 2d2, 2d12 and 2d111) in Capitella contribute their progeny to the trunk region, although the 1c, 1d, 2a, and 2c descendants are observed as scattered cells (Fig. 12) [25, 56]. Micromere 2d112 but not 2d111 in Capitella undergoes bilateral division [25]. Micromeres other than 2d and 4d in Platynereis may exclusively contribute to the head region (Fig. 12) [57].
Fig. 12 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Occurrence of micromere lineages that contribute progeny to the trunk region in Platynereis, Capitella, Tubifex and Helobdella [56,57,58]. The relationships of these taxa are based on [62]. Data for 1b are omitted here
The extent to which micromeres other than 2d and 4d contribute progeny to the trunk region in ancestral annelids is not known. In extant annelids, most, if not all, of the trunk ectoderm and mesoderm arise from two micromeres, 2d and 4d [58]. Given that these conserved features of 2d and 4d originated from the ancestral annelids, it seems likely that the trunk ectoderm and mesoderm in ancestral annelids mostly arose from 2d and 4d, as in current annelids. On this assumption, it is conceivable that the number of micromeres (other than 2d and 4d) that contribute progeny to the trunk region in ancestral annelids would have been low. At present it is equally possible that ancestral annelids lacked such micromeres except 2d and 4d. At any rate, it is most likely that more novel micromere subclones that contribute their progeny to the trunk region arose in the lineages leading to Tubifex, Helobdella, and Capitella. Novel subclones for micromeres 3c and 3d, which produce the ventral epidermis in Tubifex, fibre networks in Helobdella, and mesodermal bands in Capitella (Table 2), might have already been present in the last common ancestor of Capitella and clitellates after their evolutionary separation from Platynereis. The fate must have changed of some micromere subclones, generating novel structures.
Compared with the novel structures derived from 3c and 3d, the squamous epithelium derived from 1c and 1d in clitellate annelids (Tubifex and Helobdella) is an ordinary structure because similar epithelial sheets are seen in other annelids and even in nonannelid spiralians that undergo gastrulation by epiboly [44]. The epithelial sheets, which are the dorsal yolk sac ectoderm in Tubifex, expand across the surface to cover the entire yolky macromeres. The animal cap of micromeres in nonclitellate annelids such as Capitella and Platynereis expands via epiboly, and the resulting squamous epithelium engulfs a solid cluster of yolky macromeres [44]. The epithelial sheets derived from the micromeres in clitellates and polychaetes almost identically internalise the endoderm.
Adopting the first-quartet micromeres for epiboly appears to be unique to clitellates. The second- and third-quartet micromeres, but not those in the first quartet, in the gastropod mollusc Ilyanassa obsoleta cover endodermal clones during gastrulation [59]. Although micromere clones responsible for epiboly in polychaete embryos have not been specified, the second- and third-quartet clones likely participate in epiboly in polychaete embryos, as observed in Ilyanassa. The expansion of an animal cap (dome-shaped assemblage) of micromeres toward the vegetal pole occurs at the beginning of gastrulation similarly in polychaetes and Ilyanassa embryos [44, 59]. If this is the case for polychaetes, the mobilisation of the first-quartet micromeres for epiboly is an innovation that arose in the clitellate lineage after evolutionary separation from Capitella (Fig. 12).
Both Tubifex and Helobdella adopt the first-quartet micromere subclones for epiboly; this developmental modification may have originated from the last common ancestor of these annelids. Embryogenesis in clitellates is characterised by a large egg, large micromeres (2d and 4d), the generation of five bilateral pairs of teloblasts, and the production of well-organised GBs [15]. These large-scale developmental modifications must have originated in the last common ancestor of Tubifex and Helobdella. Thus, adopting the first-quartet micromere subclones for epiboly and these large-scale developmental modifications might have almost simultaneously occurred. This suggests that the former modification arose as a result of the latter. These large-scale developmental modifications could have led to a series of structural alterations that allowed the first-quartet micromeres to participate in epiboly during gastrulation.
The micromeres other than 2d and 4d in Tubifex embryos at the 22-cell stage organised in an arc-shaped assemblage called a micromere arc that surrounds the anterior side of the large 2d micromere. The first-quartet micromeres, especially 1c and 1d, lined up along the posterior margin of the micromere arc in contact with 2d at this stage (Fig. 1a, b) [16]. The 2d derivatives, such as the four bilateral pairs of ectoteloblasts, were located approximately 150 μm away from the posterior margin of the initial micromere arc when teloblastogenesis in micromere 2d was complete [16]. In other words, a small space existed between these structures, which was also lateral between the incipient GBs. An epithelial sheet that appeared to extend from the micromere arc already covered the underlying endoderm in this space [16]. This marked the beginning of epiboly in Tubifex embryos. Subsequently, this space and the epithelial sheet therein posterolaterally expanded as the GBs elongated towards the ventral midline on either side of the embryo (Fig. 2a, b), suggesting that the elongating GBs and the 2d derivatives that posteriorly moved created space for the first-quartet micromeres. We concluded that large-scale developmental modifications enabled the mobilisation of the first-quartet micromeres for epiboly based on these observations.
In summary, we hypothesise that these recently derived micromere subclones were mobilised for epiboly in ancestral clitellates that underwent large-scale developmental modifications. The epiboly area in Tubifex embryos (460 μm in diameter [60]) is approximately 10 times larger than that in Capitella embryos (200 μm in diameter [58]). Only two micromeres, 1c and 1d, contributed descendants to cover this epiboly area in Tubifex (Fig. 3a, b). The first quartet of micromere subclones involved in epiboly movement in the Tubifex embryo form epithelial sheets with the capacity to expand long distances and distribute to the trunk region. However, the first quartet of micromeres in polychaete embryos exclusively contribute to the head region but not to the trunk region (Table 2) [56, 57]. Novel subclones with the capacity to distribute to the trunk region likely derived from the first-quartet micromeres in ancestral clitellates or earlier. These novel micromere subclones could have already been present in the last common ancestor of Tubifex and Helobdella, which might have been readily mobilised for epiboly as the large-scale developmental modifications occurred.
Comments (0)