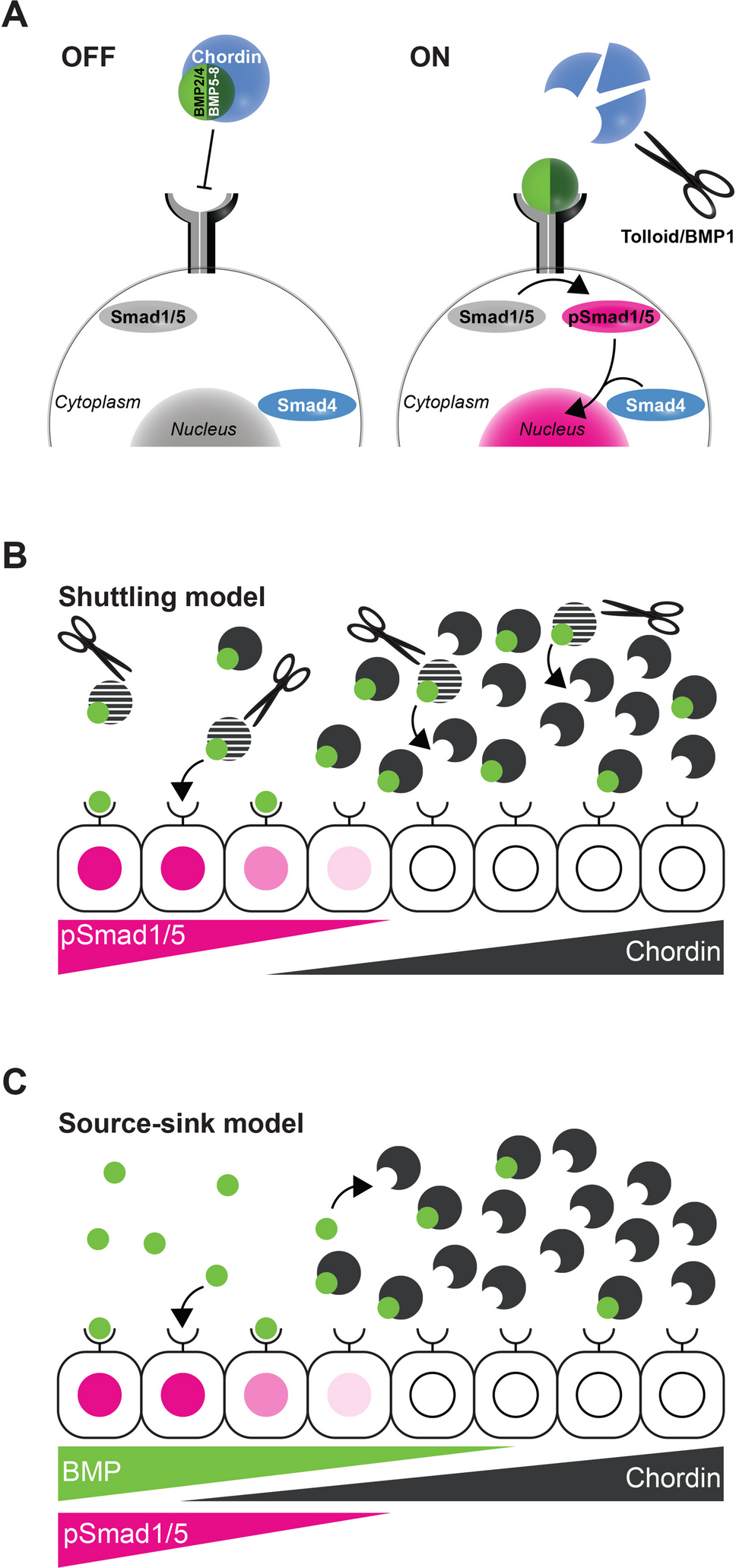
The interpretation of engrailed expression requires a clear definition of what is meant by “boundaries”. In classical developmental contexts, boundaries are typically understood as interfaces between distinct tissues or compartments, such as parasegmental boundaries, neuroepithelial compartments, or sharp interfaces between morphogenetic fields. Engrailed has historically been discussed in this framework, particularly in relation to segmentation and compartmentalization [2, 25, 26]. However, accumulating evidence from both invertebrate and vertebrate systems indicates that engrailed expression is not restricted to such tissue-level boundaries. Instead, engrailed has been reported in a variety of non-segmental contexts, including neural subpopulations [27], appendage primordia [28], and other sharply differentiated cellular domains that do not necessarily coincide with classical tissue interfaces [18, 19]. In this broader sense, “boundaries” are not defined by the presence of a multicellular tissue border, but by the establishment and maintenance of functionally and morphologically distinct cellular identities or polarized states. Such boundaries may occur at different organizational scales, ranging from interfaces between tissues to sharply defined cellular states within a single functional unit. This expanded conceptualization provides a framework for interpreting engrailed expression beyond classical segmentation-related contexts.
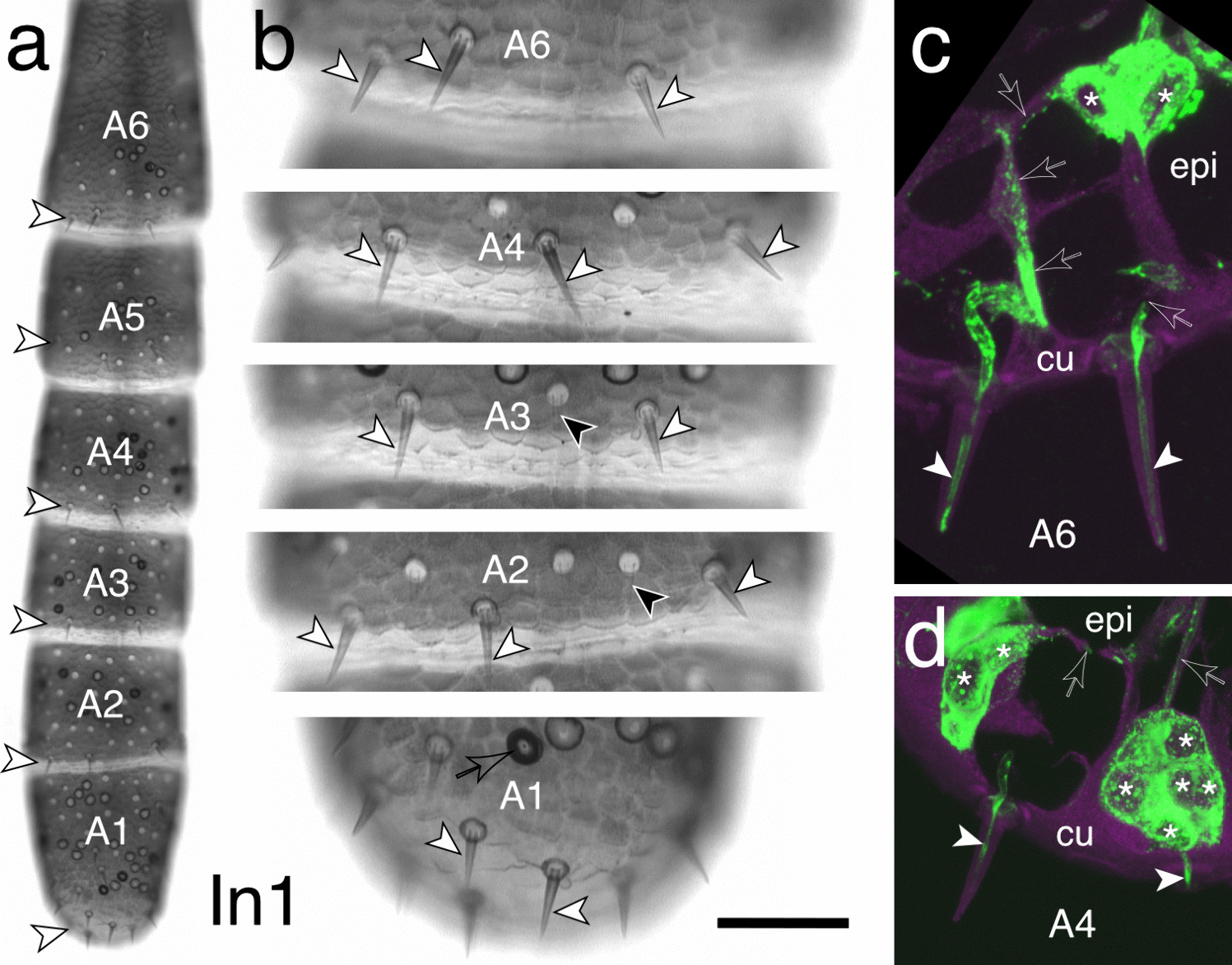
The protonephridial system of rotifers represents an extreme example of such a context. In Brachionus manjavacas, this system does not constitute a tissue in the conventional sense, but rather a minimal functional unit composed of a small number of highly specialized and polarized cells, including terminal and canal cells [14]. Importantly, engrailed expression in excretory or protonephridial organs has not previously been reported from other metazoan taxa. However, members of other conserved developmental gene families have been implicated in excretory system formation across Bilateria. Pax2/5/8 genes are expressed in nephric or protonephridial systems in vertebrates (e.g., Mus musculus) [29], annelids (Helobdella austinensis) [30], onychophorans (Euperipatoides rowelli) [31], and molluscs [32], consistent with roles in excretory tissue specification. In vertebrates, Pax2/5/8 function together with Six and Eya family members during early kidney development as part of a conserved regulatory network [33]; likewise, orthologous Six1/2 and Eya genes, together with Osr, POU2/3, Sall, and FoxQD, contribute to protonephridial regeneration and differentiation in the planarian Schmidtea mediterranea [34]. FoxD1 further represents a classical component of vertebrate kidney patterning [35]. Against this broader background, the deployment of engrailed in the rotifer protonephridial system represents a novel addition to the repertoire of conserved patterning genes associated with excretory structures. I therefore do not interpret the observed pattern as evidence for a conserved excretory role of engrailed. Instead, I suggest that engrailed has been lineage-specifically recruited in rotifers to stabilize sharply defined cellular states within a highly miniaturized functional system. In this view, the protonephridial engrailed expression in rotifers reflects neither segmentation nor organ homology, but rather the deployment of a conserved regulatory factor in a novel cellular context characterized by extreme polarity and functional differentiation.
The expression pattern of engrailed in Brachionus manjavacas adds an unexpected dimension to the already remarkable diversity of en deployments across metazoan development. The most pronounced expression territory within the terminal and canal cells of the protonephridial system differs fundamentally from any pattern previously described in bilaterian or non-bilaterian animals. Rotifer protonephridia consist of very few cells, yet they possess hallmark epithelial features, including pronounced apico–basal polarity, intercellular junctions, and highly differentiated ciliary structures [14, 36]. These characteristics define them as microanatomical units with polarized architecture despite their extreme cellular reduction. Against this background, the presence of engrailed in terminal and canal cells is consistent with a role in the organization or stabilization of polarity domains and the coordination of ciliary differentiation within a compact excretory structure. In this context, engrailed operates at the level of individual cells or small cell clusters rather than at macroscopic tissue-level boundaries such as parasegment borders, shell fields, or apical–mantle margins [3, 4, 37]. This small-scale deployment resonates with the widely discussed view that the ancestral function of engrailed was not segmentation, but the stabilization of sharply defined developmental interfaces [4, 38], as outlined above. Across Metazoa, engrailed is repeatedly associated with regions where abrupt changes in cell identity, polarity, or tissue organization must be maintained (see Table 1 for an overview). The rotifer data extend this conceptual framework by demonstrating that engrailed can also be deployed in highly reduced, unicellular or bicellular structures where polarity, ciliary function, and directional flow converge.
The second engrailed domain in B. manjavacas, a signal in neurons associated with the developing foot, reinforces this interpretation. Rotifer pedal adhesion supposedly requires sensory processing, secretory control, and coordinated behavior, and the neurons in the foot represent a hub where epithelial, neuronal, and secretory functions meet [14]. Engrailed expression in these cells therefore points to a preserved neurogenic potential. Across bilaterians, engrailed is expressed in diverse neuronal subsets, including interneurons of the vertebrate spinal cord, components of the insect CNS, and elements of the nervous systems in some spiralian lineages [7, 39,40,41]. The rotifer pattern is congruent with this broader distribution, despite the pronounced miniaturization and reduced cellular complexity characteristic of many gnathiferan lineages.
The protonephridial and pedal-neuronal expression domains represent functionally independent territories, yet both are sharply defined and highly specialized. Their shared feature is not epithelial architecture, but a requirement for stable cellular identity and polarized, directional activity - ciliary transport in the terminal organ and sensory–secretory integration in the foot. The presence of engrailed in such distinct, precisely delimited cell types supports the notion that the gene is recruited to developmental contexts where discrete functional boundaries or transitions between polarized cellular states must be maintained, rather than to broad epithelial sheets or segmentation modules.
It may be argued that boundary maintenance is unnecessary in structures composed of only one or two cells, where the plasma membrane itself defines the physical limit of the cell. Moreover, transcription factors are frequently co-opted for lineage-specific functions during evolution, and it remains possible that engrailed has acquired additional or modified roles in rotifers. However, even unicellular or bicellular systems require the establishment and stabilization of sharply polarized subcellular domains and functional compartments. In highly specialized cells such as protonephridial terminal organs, apico–basal polarity, ciliary apparatus organization, and directional transport depend on precisely regulated intracellular patterning. In this context, “boundary” does not refer to a multicellular tissue interface, but to the maintenance of discrete functional domains within or between specialized cells.
Importantly, the rotifer pattern provides an independent test of longstanding hypotheses about the origin of segmentation [42, 43]. I note that the conclusions drawn here are based on the expression pattern of a single gene and do not, by themselves, constitute evidence for segmentation; broader assessments would require data on additional components of the segment polarity gene network. If the ancestral function of engrailed involved the maintenance of local epithelial discontinuities, then segmental stripes in annelids and arthropods are best viewed as a lineage-specific elaboration of this boundary program rather than as evidence that segmentation is an ancient bilaterian trait [43, 44]. The absence of any segment-like deployment in rotifers, combined with their unique expression territory in a protonephridial organ, strengthens the interpretation that large-scale morphological segmentation evolved independently in annelids and arthropods through the repeated co-option of older, more flexible patterning systems [44].
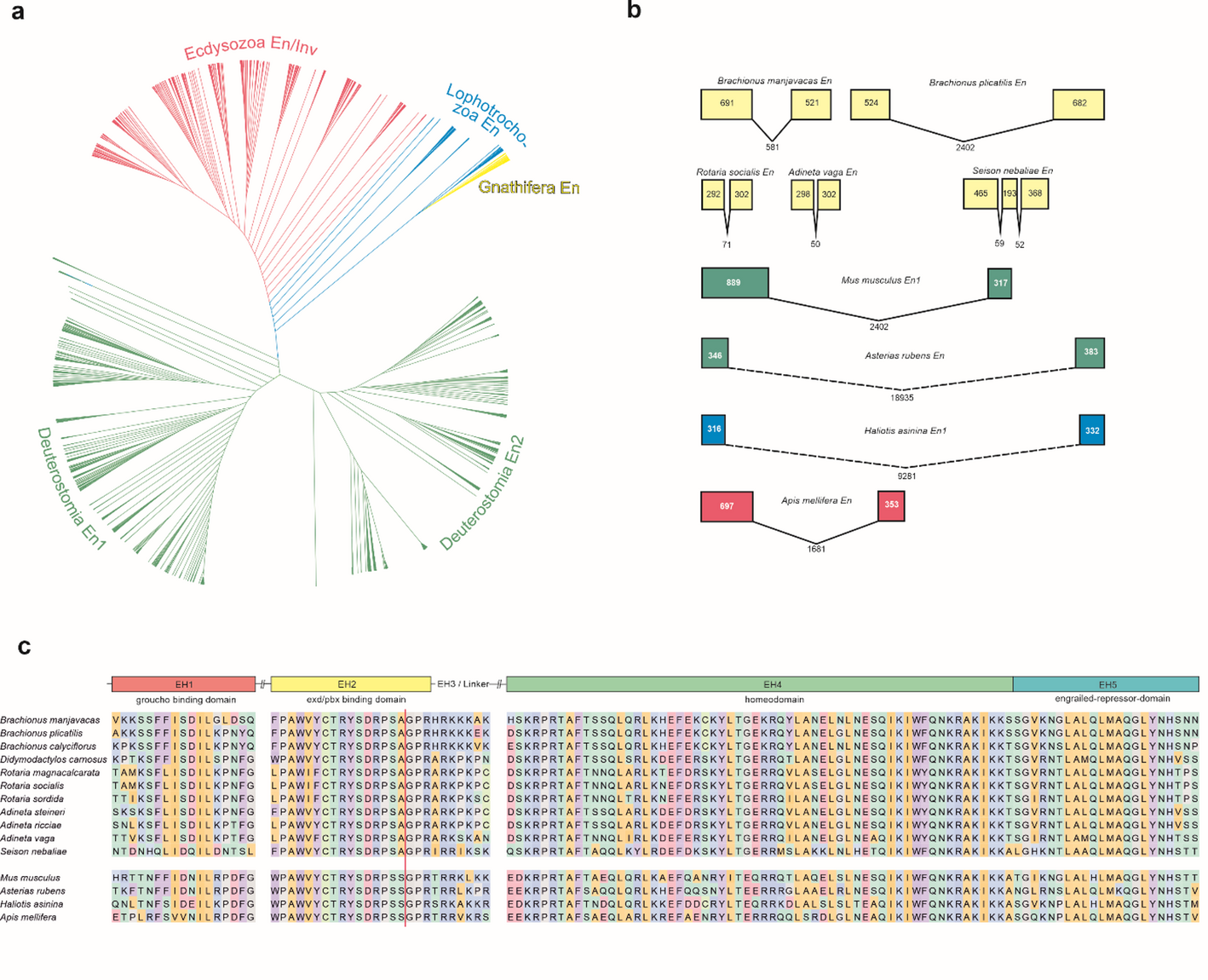
In addition to its expression pattern, the genomic architecture of engrailed provides further insight into its evolutionary stability. The EH2 intron identified in Brachionus manjavacas occupies an invariant position and phase relative to the coding sequence across protostomes and deuterostomes. Conservation of intron positions is a well-documented phenomenon in eukaryotic evolution and is frequently associated with orthologous relationships and structural constraint on gene architecture [45,46,47]. At the same time, extensive lineage-specific intron loss and gain have shaped intron landscapes across Metazoa. In this broader context, the metazoan-wide conservation of both position and phase of the EH2 intron is particularly striking. While intron positional conservation is not unique to engrailed, the metazoan-wide stability of this intron within a deeply conserved regulatory gene underscores strong structural constraint at this locus.
Finally, rotifers highlight how conserved regulatory genes can be integrated into extremely reduced developmental systems without losing their fundamental logic. Engrailed is retained where polarity must be enforced, where ciliary architecture requires coordination, and where sensory–secretory functions converge. This underscores the robustness of engrailed as a developmental regulator across dramatic variation in body size, organ complexity, and cell number. Gnathiferan taxa therefore offer an independent and previously underrepresented perspective on bilaterian patterning mechanisms and highlight the importance of microanatomical model systems for understanding the evolutionary trajectories of conserved developmental regulators.


Comments (0)