
Remember me
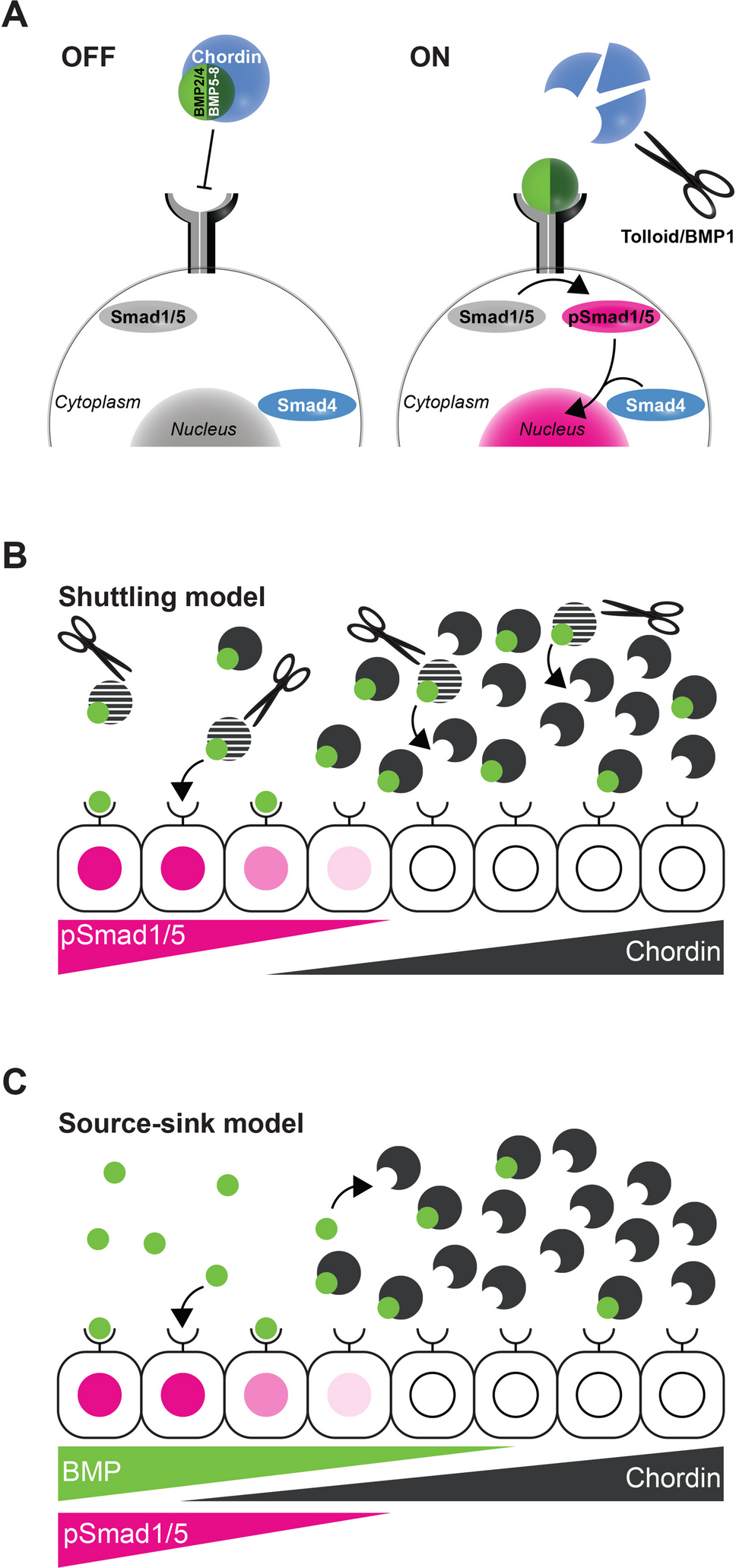
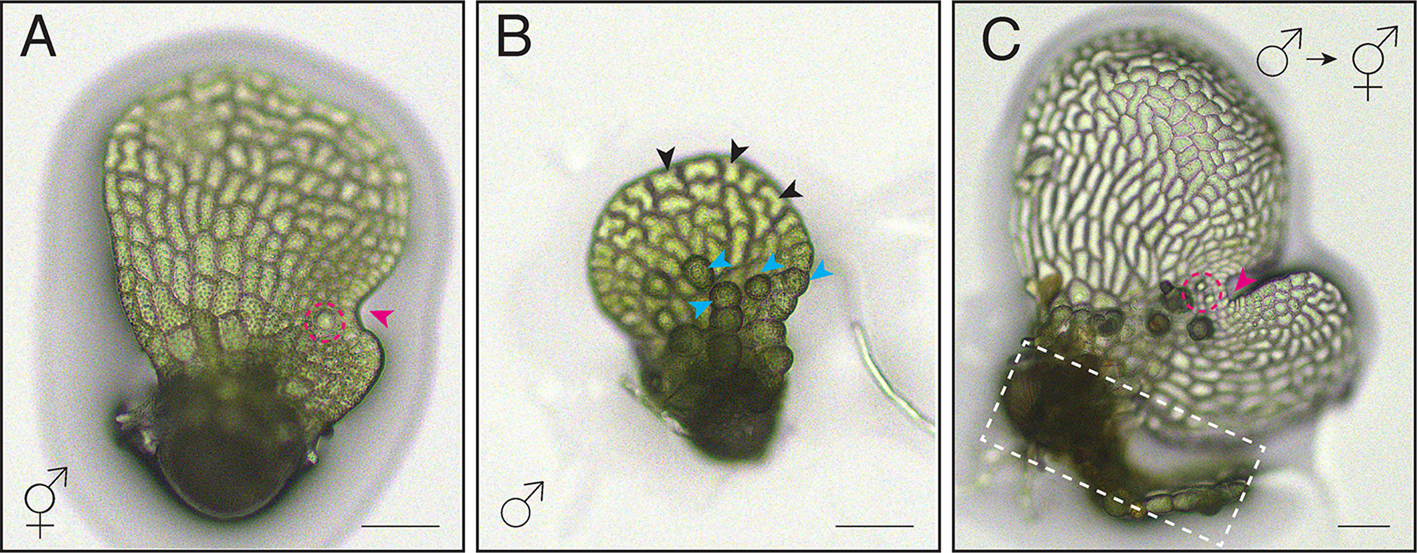

Under the influence of the pheromone antheridiogen, Ceratopteris gametophytes germinating from genetically identical spores exhibit sexual dimorphism, developing either as males or hermaphrodites [6, 23] (Figs. 1A-C and 2). Ceratopteris haploid spores that germinate in an antheridiogen-free environment, or that germinate early within a population before antheridiogen accumulates, develop as hermaphrodites (Figs. 1, 2, 3 and 4). They initiate a multicellular marginal meristem and subsequently form egg-producing archegonia adjacent to the meristem (Figs. 1A, 2 and 4). Once initiated, the hermaphrodite developmental program is generally irreversible by antheridiogen, as the marginal meristem remains indeterminate, sustains cell proliferation, and induces archegonium formation. Meanwhile, hermaphrodites can also produce a few sperm-producing antheridia outside the meristem, typically at the edges of the prothallus [6, 12, 24]. Once specified, Ceratopteris hermaphrodites begin producing and releasing antheridiogen into the surrounding environment [6, 23], which induces late-germinating, undetermined gametophytes within the population to develop as males. Unlike hermaphrodites, males are ameristic, lacking any meristem or archegonia, but gradually develop a number of sperm-producing antheridia (Figs. 1B, 3 and 4). Upon maturation, antheridia rupture and release motile, flagellated sperm, which swim into the archegonia of hermaphrodites to fertilize eggs and producing diploid embryos, advancing the life cycle [11, 12, 23]. Through this process, antheridiogen adjusts sex ratios in the Ceratopteris gametophyte population and facilitate sexual reproduction.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Sex types in Ceratopteris Gametophytes. (A-C) Light micrographs showing a Ceratopteris hermaphrodite (A), a male (B), and male-to-hermaphrodite conversion (C). Magenta arrowheads: meristem in (A) and de novo meristem in (C); magenta dashed circles in (A, C): archegonia; blue arrowheads in (B): representative antheridia; black arrowheads in (B): representative non-antheridium cells; white-dashed box in (C): original male body from which the de novo meristem forms. Scale bars in (A-C): 100 μm
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.The multicellular meristem and adjacent archegonia in Ceratopteris hermaphrodites. A hermaphrodite (5 days after germination) stained and imaged by confocal microscopy. The hermaphrodite develops a multicellular meristem on one lateral side of the prothallus, characterized by small cells forming a concave notch. Yellow bar: meristem notch region; white dash circle: developing archegonium adjacent to the meristem. Magenta: cell outlines. Scale bar: 50 μm
Fig. 3 The alternative text for this image may have been generated using AI.
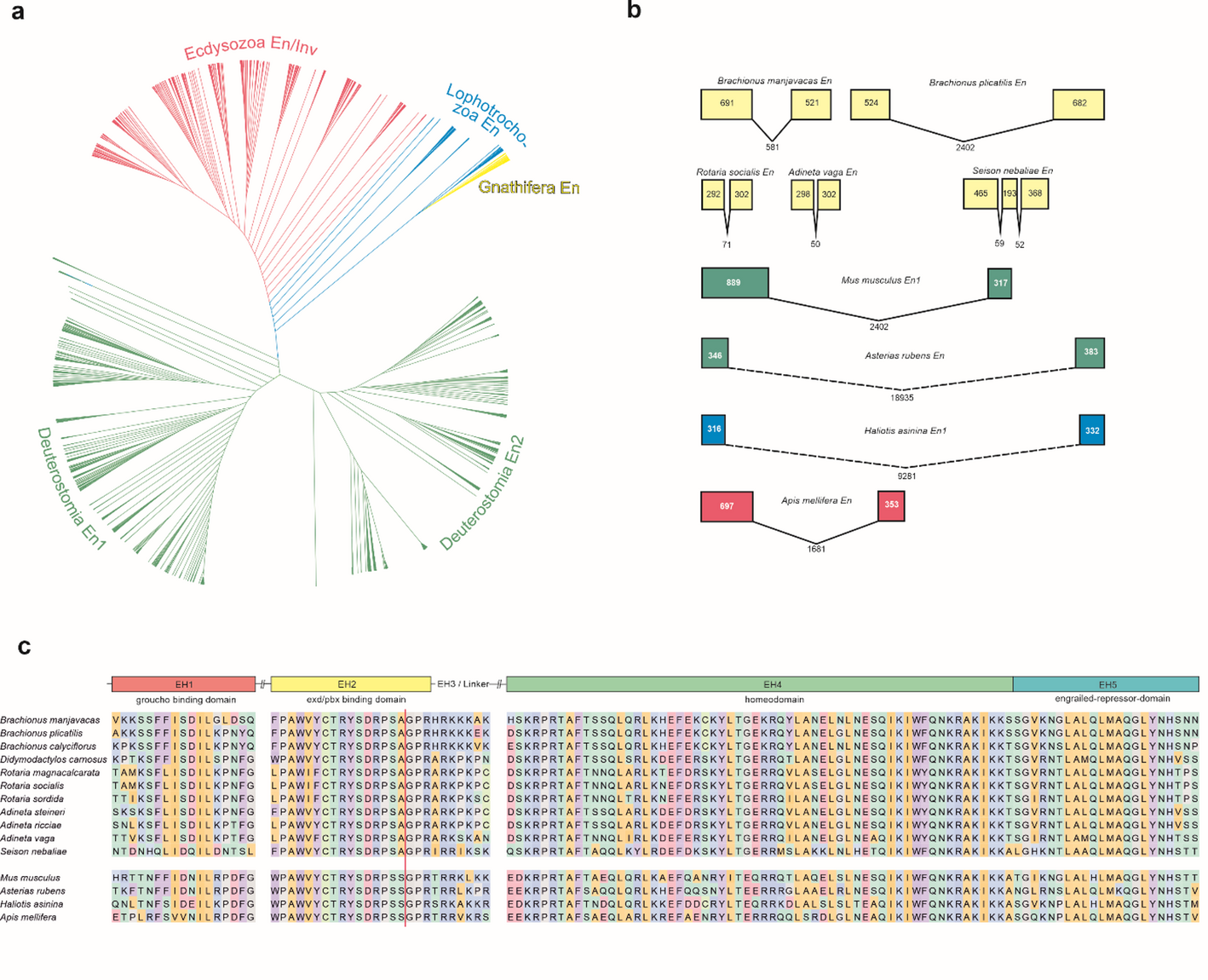
The alternative text for this image may have been generated using AI.Sex-type specification pathways in Ceratopteris gametophytes. (A-C) Pie charts illustrating sex ratios in Ceratopteris gametophyte populations of different genetic backgrounds, indicating the proportion of hermaphrodites (pink) and males (blue). (A) Ceratopteris wild type (WT). (B) her7 mutants (adapted from Banks, 1994) (C) CrHAM loss-of-function and miR171 gain-of-function lines (adapted from Geng et al., 2024). (D) Diagram illustration of the miR171-CrHAM regulatory module in Ceratopteris gametophytes. CrHAM promoter activity and CrHAM protein accumulation are illustrated on a scale from white (none) to red (high). The figure was created using BioRender and further modified in PowerPoint for clarity
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Diagram illustrating Ceratopteris sex determination and sex-type conversion. In WT, spores germinate and develop into hermaphrodites when not exposed to antheridiogen (Ace), developing a multicellular meristem and adjacent archegonia. When exposed to antheridiogen, spores instead germinate and develop into males, which lack both a meristem and archegonia but produce sperm-bearing antheridia. In the continued presence of antheridiogen, males maintain their identity and produce additional antheridia. In contrast, removal of antheridiogen triggers male-to-hermaphrodite conversion, and a meristem-progenitor cell (MPC) is established within the male body, and its progeny give rise to the entire de novo meristem. In her7 mutants, spores develop into hermaphrodites in both antheridiogen-free and antheridiogen-present conditions. The figure was created using BioRender and further modified in PowerPoint for clarity
Antheridiogens from several ferns have been identified as gibberellin-related compounds [25,26,27]. For example, in L. japonicum, the antheridiogen shares a common ent-gibberellane skeleton with gibberellin A4 (GA4), but differs structurally by lacking an OH group at C3 and possessing a methyl-esterified carboxyl group at C6 [25, 27, 28]. In contrast, the chemical structure of the antheridiogen in Ceratopteris richardii remains unknown. Nevertheless, it has been proposed that Ceratopteris antheridiogen signaling likely, or at least in part, involves the GA pathway [10, 29] based on previous experimental observations. For instance, treatment of GA biosynthetic inhibitors reduced the proportion of males in Ceratopteris gametophyte populations, suggesting that antheridiogen and GA share similar or overlapping biosynthesis steps in Ceratopteris [29]. Additionally, abscisic acid (ABA) antagonizes antheridiogen actions, inhibiting antheridiogen-induced antheridium formation and male development in Ceratopteris gametophytes [10]. Several Ceratopteris mutants have been isolated that are insensitive to ABA treatment, enabling the antheridiogen-triggered male differentiation programs to proceed even in the presence of ABA [30,31,32]. Among them, one mutated gene has been identified as GAIA1, which encodes a close homolog of Arabidopsis OST1, a conserved SnRK2 signaling component [32]. Despite these advances, the cellular basis of ABA-mediated cell fate specification and cell proliferation during fern gametophyte development remains to be uncovered. Additionally, in seed plants, antagonism between gibberellin and ABA regulates various developmental progresses, including seed dormancy and germination, root growth, and response to environmental stimuli [33]. Dissecting the molecular mechanisms that mediate the antagonistic interaction between antheridiogen and ABA during Ceratopteris sex determination will be an exciting direction for future research.
Forward genetic screens have identified several mutants affecting sex type in Ceratopteris gametophytes, including hermaphroditic (her), transformer (tra), and feminization (fem) mutants [6, 11, 31, 34]. her gametophytes are insensitive to antheridiogen, and upon germination, they develop into hermaphrodites even in the presence of antheridiogen [31] (Figs. 3A-B and 4). The tra mutation disrupts both meristem and archegonia formation, causing gametophytes to differentiate into ameristic males even under conditions that normally promote hermaphrodite development [31]. In contrast, feminization (fem) mutants fail to form antheridia and instead promote meristic female development in gametophytes [31]. Recently, her7 mutations were identified in a brassinosteroid receptor-like kinase homolog in Ceratopteris [35]. Bulk segregation and sequencing revealed that five independent her7 alleles (her7-1, her7-11, her7-14, her7-15, and her7-19) localized to the same locus within the mapping window on chromosome 29, each carrying either missense mutations or small amino acids deletions in this receptor-like kinase homolog [35]. HER7 localizes to the plasma membrane and cytoplasm, consistent with its proposed role as a receptor kinase [35]. Transcriptomic analyses comparing wild type and her7 mutants at 12 days post inoculation, as well as young gametophytes (4.5 days post inoculation) treated with or without antheridiogen, revealed global gene expression changes in response to antheridiogen, many of which appear to be dependent on HER7 [35, 36]. Specifically, differential expression of genes involved in brassinosteroid (BR) and GA biosynthesis and signaling provides important molecular clues for antheridiogen sensing and points to promising directions for future studies.
2.1 Maintenance of hermaphrodite identity and meristem activityAs described above, a key regulatory mechanism ensuring the stability of this system is that, though antheridiogen induces male developmental programming, hermaphrodites, once specified, become insensitive to antheridiogen produced either by themselves or neighboring individuals. Without this insensitivity, hermaphrodites would not be able to maintain their sex type, leading to male dominance across the population. A central outstanding question in the field is: what molecular mechanism orchestrate meristem identity maintenance and antheridiogen insensitivity in hermaphrodites? Recent studies have identified a HAIRY MERISTEM (HAM) family GRAS-domain transcriptional regulator as a key player in this process [20, 37]. HAM proteins are widely present across plant lineages and are evolutionarily conserved in land plants [38, 39]. In seed plants such as Arabidopsis, the HAM family play essential roles in maintaining stem cell indeterminacy and proliferation within shoot apical meristems [39,40,41,42,43,44]. In Ceratopteris, the HAM homolog (CrHAM) was shown to suppress antheridiogen-mediated male differentiation while maintaining meristem indeterminacy and cell proliferation in hermaphrodites, thereby contributing to the regulation of sex ratios in gametophyte populations [20].
CrHAM protein preferentially accumulates in the meristems of hermaphrodites but is excluded from antheridia in males [20]. CrHAM knockdown (KD) in Ceratopteris transgenic lines increased the male ratio in gametophyte populations (Fig. 3C) and impaired hermaphrodite development, resulting in smaller prothalli, reduced meristem size, and a shallower meristem notch compared with wild type (WT). In many KD hermaphrodites, archegonia were absent or greatly reduced near abnormal meristems, whereas antheridia formed ectopically around and even within the meristem. Consistently, male-specific GAMYB genes were up-regulated in CrHAM KD gametophytes. In antheridiogen treatment assays, WT hermaphrodites retained their sex identity, but a considerable number of young CrHAM KD hermaphrodites failed to maintain meristem activity and gradually converted into ameristic males, explaining the elevated male ratio in KD populations. These results suggest that CrHAM is required to maintain meristems in an undifferentiated state when exposed to antheridiogen. In addition to suppressing male traits, CrHAM promotes female traits by sustaining cell proliferation during hermaphrodite development [20], as CrHAM KD exhibited reduced prothallus size and cell number due to decreased division activity. Transcriptomic analyses further reflected the positive roles of CrHAM in promoting meristem activity and hermaphrodite development. Gene Ontology (GO) analysis of differentially expressed genes (DEGs) in CrHAM KD hermaphrodites indicate CrHAM involvement in cell growth and prothallus expansion [20]. Additionally, a CLAVATA1 homolog (CrCLV1a) and an auxin biosynthesis gene, the YUCCA homolog (CrYUC1) were downregulated in CrHAM KD hermaphrodites. Both genes are generally expressed at higher levels in WT hermaphrodites compared with WT males [20]. This finding was consistent with studies in the liverwort Marchantia, where MpCLV1 and MpYUC2 are expressed in the meristem and required for cell proliferation in the notch [45, 46].
The transcriptional reporter showed that the CrHAM promoter is active in nearly all cells of hermaphrodite prothalli (except gametes), whereas the translational reporter revealed CrHAM protein enrichment specifically at the meristem [20] (Fig. 3D). This raises an important question: what factors control the spatial restriction of CrHAM protein accumulation? In Arabidopsis, type II HAM genes (AtHAM1, AtHAM2 and AtHAM3) are negatively regulated by microRNAR171(miR171) [38, 39, 47, 48]. In Arabidopsis SAMs, MIR171/170 are specifically expressed in the epidermis and move downwards within a limited distance into deeper layers, and they mediate the cleavage of AtHAM1-3 transcripts, generating an apical–basal concentration gradient of HAM proteins [49, 50]. Interestingly, the CrHAM coding sequence contains a miR171-binding site (5’-GATATTGGCGCGGCTCAATCA-3’) identical to that in AtHAM1-3 genes, indicating a conserved regulatory mechanism [39]. Supporting this, two Ceratopteris MIR171 genes (CrMIR171B and CrMIR171C) have been identified, and their mature miRNAs share nearly identical sequences with Arabidopsis miR171b/c [20]. Overexpression of CrMIR171B in Ceratopteris reduced CrHAM expression and produced defects resembling CrHAM KD, including an increased male ratio in gametophyte populations (Fig. 3C, D), and smaller prothalli and shallower meristem notches in hermaphrodites [20]. Together, these findings provide evidence that the miR171 family is present and functional in Ceratopteris, where it contributes to sex determination and meristem maintenance likely through negatively regulating CrHAM expression [20, 37, 39]. This highlights a conserved yet lineage-specific miR171-HAM regulatory module in ferns, opening promising directions for future studies.
2.2 Male-to-hermaphrodite conversion and de novo meristem formationCeratopteris gametophytes exhibit remarkable sexual developmental plasticity, with sex types capable of switching in response to environmental cues (Fig. 4). Previous studies demonstrate that Ceratopteris hermaphrodites continuously produce and secret antheridiogen, which not only induces sexually undetermined gametophytes to develop as males but is also required for maintaining the male developmental program [12]. When antheridiogen is removed or depleted, male identity cannot be maintained, and male gametophytes initiate de novo meristems and produce egg-bearing archegonia, thereby converting into hermaphrodites [12, 22, 23](Figs. 1C and 4). This environmentally responsive strategy allows ferns to adjust population sex ratios under changing conditions, promoting outcrossing and enhancing reproductive success. This naturally occurring male-to-hermaphrodite conversion provides an ideal model for dissecting the molecular and cellular mechanisms of cell fate specification and the coordination of cell division during de novo meristem formation. Recent work has combined long-term, non-invasive time-lapse confocal imaging with computational image analysis of a ubiquitously expressed fluorescence nuclear marker, capturing the complete developmental trajectory of this process at high spatiotemporal resolution [22].
This study reconstructed the dynamic lineage maps of sex-type conversion and revealed that the entire newly formed multicellular meristem originates from a single non-antheridium cell, the meristem progenitor cell (MPC), in the male prothallus [22] (Fig. 4). Over time, the MPC lineage expands and sustains active cell division, in contrast to other cell lineages, which gradually become mitotically inactive during meristem formation and sex-type conversion. In parallel, a mathematical model demonstrated that a combination of stochastic cell division and inhibitory signals from actively dividing cells is sufficient to account for MPC lineage establishment and meristem formation [22]. Furthermore, treatment with Aphidicolin, a drug that specifically inhibits the S phase of the cell cycle, disrupted male-to-hermaphrodite conversion and led to the reduced size of de novo meristems [22], consistent with the simulation generated using adjusted parameters for reduced cell division. These findings demonstrated that re-entry into and maintenance of cell cycle progression are essential for male-to-hermaphrodite conversion and MPC lineage establishment [22].
Division orientation and cell position further influence division activity during de novo meristem formation. In particular, the orientation of the initial division in the MPC determines the proliferative activity of its progeny: an anticlinal division produces two daughter cells in the outermost layer, giving rise to two lineages with similarly high division activity, whereas a periclinal division generates one daughter cell in the outermost layer, which maintains higher division activity in its lineage, and another in the inner layer, which produces a cell population with lower activity [22]. Quantitative analyses show that, once de novo meristems are established, cells in the outermost layer divide significantly more frequently than those in the inner layers [22]. This pattern is consistent with previous findings that the outermost (marginal) layer of meristem cells in Ceratopteris hermaphrodites exhibits more active division activity compared with inner cells [17]. Interestingly, recent work in the epiphytic fern Phlebodium pseudoaureum (Polypodiaceae) also demonstrated that the outermost layer predominantly drives active cell division during de novo meristem formation and notch establishment [51]. Together, these results suggest that in Ceratopteris gametophytes, meristem development during normal hermaphrodite growth and de novo meristem formation during sex-type conversion share conserved strategies, driven by positional signals from the outermost layer, to sustain cell proliferation. These strategies also appear to be conserved across fern species from diverse taxa.
Comments (0)