
Remember me
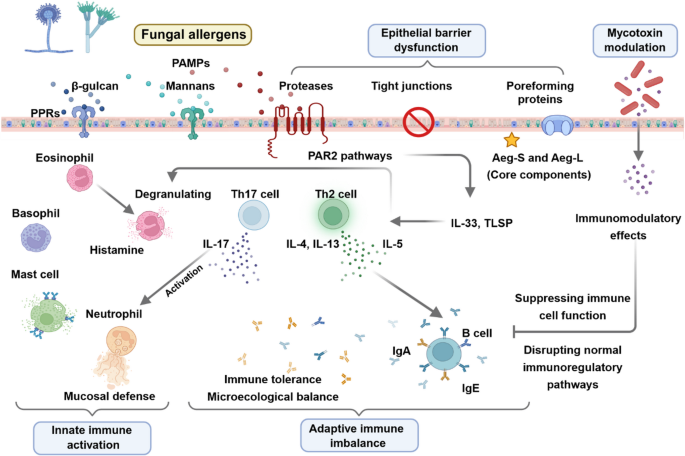
Asthma is an inflammatory disease affecting approximately 300 million people worldwide and is characterized by airway hyperresponsiveness, mucus overproduction, and airway remodeling. Its pathogenesis is driven by complex interactions between epithelial and immune cells, including Th2 lymphocytes, dendritic cells (DCs), mast cells, and eosinophils, which together orchestrate type 2 inflammation [19]. Airway remodeling observed in the course of asthma is a complex process that involves structural, cellular, and molecular alterations. One of them is thickening of the subepithelial extracellular matrix (ECM) with increased collagen deposition (collagens I, II, and VI) accompanied by goblet cell hyperplasia and smooth muscle hypertrophy [20]. Furthermore, complex interactions between epithelial cells, fibroblasts, and mast cells within proinflammatory hubs occur in spatially-organized niches that produce alarmins and chemokines [21]. One of the most critical remodeling factor is epithelial cell-derived lumican acting by promoting fibroblast proliferation and collagen production while the balance between matrix metalloproteinases and tissue inhibitors determines whether remodeling proceeds through ECM deposition in acute models or ECM degradation in chronic inflammation models. Airway remodeling is also characterized by apoptosis of infiltrating immune cells in acute models localized to alveolar regions, and by necroptosis in structural cells during chronic remodeling, which releases pro-inflammatory mediators and accelerates innate immune responses [22]. Bronchoalveolar lavage fluid (BALF) remains a gold standard for assessing airway inflammation, enabling analysis of airway and alveolar components and provides valuable insights into disease mechanisms through proteomic and genomic analyses [23].
Altered EVs profiles observed in BALF from asthmatic patients compared to healthy controls suggest that they actively contribute to airway inflammation and asthma pathogenesis rather then merely reflecting it [24]. One of the key mechanisms by which EVs modulate asthma pathogenesis is the transfer of microRNAs, which regulate gene expression in recipient cells primarily through mRNA degradation or translational inhibition [25]. They are involved in numerous biological processes such as lung development, immune responses, and the pathogenesis of pulmonary diseases such as lung cancer, asthma, chronic obstructive pulmonary disease (COPD), and pulmonary fibrosis [26]. The functions of extracellular vesicle miRNAs in allergic diseases are summarized in Table 1.
Table 1 Role of extracellular vesicle miRNAs in allergic diseasesThe presence of miRNAs within airway-derived EVs was first demonstrated by Sinha et al., who identified exosome-enclosed miRNAs in exhaled breath condensate (EBC) from asthmatic patients, highlighting their diagnostic potential [45, 46]. Since then, numerous miRNAs have been linked to asthma pathogenesis. For instance, miRNA-125b-5p has been associated with inflammatory responses in asthma, with lower expression in bronchial biopsy correlating with more severe airflow obstruction in asthmatic patients [47]. This miRNA was also identified in serum-derived exosomes from patients with asthma, where its expression was elevated and positively correlated with disease severity [27] (Table 1; Fig. 1). Similar observations were reported by Atashbasteh et al. who demonstrated increased expression of miR-125b in the plasma exosomal fraction of patients with severe asthma which correlated with inflammatory indices measured in serum, including IgE and hs-CRP levels [48]. The occurrence of opposite correlations between allergic disease severity and miRNA levels in EVs isolated from serum and tissues may be explained by several biological mechanisms. First, cells are able to selectively package specific miRNAs into EVs and release them into circulation, resulting in increased EV miRNA levels in serum and decreased EV miRNA levels in extracellular environment. Second, the export of regulatory miRNAs via EVs may serve as a mechanism to reduce their intracellular concentration or to facilitate intercellular communication by modulating signaling pathways in recipient cells. Finally, allergic inflammation may alter EV biogenesis and cargo selection, thereby influencing the enrichment of specific miRNAs in EVs and leading to disease-specific expression patterns [49, 50]. Metabolomic and cytokine profiling studies further support the concept that local airway alterations are reflected systemically. Changes observed in BALF and lung tissue, including metabolic dysregulation and cytokine shifts, are often mirrored in serum, indicating that EV-associated signals can propagate beyond the lung environment [51]. Interestingly, Hogea et al. showed correlation between the levels of cytokines in serum and BALF in patients with lung cancer and chronic lung disease [52]. Cytokine levels were significantly elevated in patients with malignancies, and analysis of BALF showed higher concentrations than in serum. Notably, cancer-associated cytokines appeared earlier and at greater levels in lavage fluid than in peripheral blood. This clearly shows that changes observed in BALF or lung tissue are frequently reflected in the serum, indicating that local pulmonary alterations, including shifts in EVs profile, can manifest systemically. Therefore, EVs obtained noninvasively from nasal lavage or serum may serve as a minimally invasive “liquid biopsy,” providing valuable information on disease status and progression through biomarker analysis.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.EVs derived from asthmatic patients contribute to the progression of the disease. Reduced levels of miR-34a, miR-92b, and miR-210 in extracellular vesicles isolated from nasal lavages of children with asthma, compared with healthy controls, are associated with airway obstruction [33]. EVs isolated from serum also differ in miRNAs content in asthmatics and healthy donors. The levels of miRNA-125b-5p, miRNA-126 and miR-122-5p molecules are elevated and correlate with more severe airflow obstruction in asthmatic patients [27,28,29]. Eosinophils modulate their own activity via the secretion of extracellular vesicles (EVs). EVs isolated from blood eosinophils of patients with asthma increase the production of reactive oxygen species (ROS) and nitric oxide (NO), enhance adhesion to fibronectin, and promote eosinophil survival. In addition, these EVs induce apoptosis in human small airway epithelial cells (SAEC) and delay wound repair by modulating the phosphoinositide 3-kinase/protein kinase B(PI3K/AKT) and Janus kinase–signal transducer and activator of transcription (JAK-STAT) signaling pathways [53, 54]
Experimental models further support the role of EV-associated miRNAs in pulmonary inflammation. Qiao et al. demonstrated dysregulation of several miRNAs in serum exosomes from rats with pulmonary neutrophilic inflammation induced by zinc oxide nanoparticles, suggesting specific miRNAs as mediators of pulmonary inflammation [31]. Among these, miR-134-5p, miR-207, miR-465-5p, miR-30b-5p, miR-19a-3p, and miR-130a-3p were identified as the most likely contributors to pulmonary inflammation in this model (Table 1; Fig. 2).
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Animal models used to elucidate the contribution of extracellular vesicles (EVs) to airway allergic inflammation. Pulmonary neutrophilic inflammation induced by zinc oxide nanoparticles in rats resulted in increased levels of miR-134-5p, miR-207, miR-465-5p, miR-30b-5p, miR-19a-3p, and miR-130a-3p in serum extracellular vesicles compared with control animals, highlighting their potential role in the development of airway inflammation in this model [31]. In a mouse model of ovalbumin (OVA)-induced asthma, bronchial epithelial cells (BEC) and macrophages display enhanced extracellular vesicle (EV) secretory activity compared with other cell types, as determined by immunohistochemical analysis. These EVs are proposed to stimulate macrophage proliferation and chemotaxis, thereby intensifying the inflammatory response [55]. Dietary consumption of bovine milk may influence pulmonary inflammatory responses to agricultural dust (DE) exposure. Mice receiving the diet with EVs disrupted by sonication (DEV) had an increased macrophage influx into lungs and decreased levels of IL-6, CXCL1, and amphiregulin (AREG) in the BAL compared to those fed the intact EV diet. This may suggest that acute lung inflammatory response elicited by DE may be modulated, at least in part, by the EVs cargo present in the diet. Ex vivo stimulation with DE revealed that macrophages from the EV-fed group produced higher levels of M1-associated proinflammatory cytokines, including TNF-α, IL-12/23, IL-6, while levels of M2-associated protective mediators such as IL-10, and arginase were reduced indicating that milk-derived EVs regulate inflammation through effects on macrophage polarization [56]
Consistent with these findings, elevated levels of miRNA-126 were detected in serum exosomes from asthmatic patients as well as in lung tissues from asthmatic mice, further supporting its involvement in asthma-related disease mechanisms [28] (Table 1; Fig. 1). In vitro studies demonstrate that epithelial cell-derived EVs actively shape immune responses; for example, IL-13-stimulated bronchial epithelial cells release EVs with altered miRNA profiles. An RT-qPCR‐based profiling of the EV‐associated miRNAs revealed decreased levels of miR‐92b, miR‐34a, and miR‐210 in IL-13-treated cells [33] (Table 1; Fig. 3).
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Involvement of EVs isolated from cell cultures in asthma pathogenesis. Human bronchial epithelial cells (HBEC) isolated from asthmatic patients produce apically EVs participating in the mammalian target of rapamycin (mTOR) and mitogen-activated protein kinase (MAPK) signalling pathways while basolaterally secreted EVs are enriched in miRNAs from the miR-320, miR-181, miR-550, let-7, and the miR-154 families involved in the T and B cell receptors signalling pathways [34]. IL-13-treated HBEC secrete EVs with decreased levels of miR-92b, miR‐34a, and miR‐210 associated with airways obstruction probably resulted from the dendritic cell maturation and development of the Th2 response [33]. EVs derived from PM2.5- treated HBE cells are enriched in miRNA let-7i-5p which inhibits the DUSP1 gene which is a target in the activation of the MAPK signalling pathway leading to the bronchoconstriction in mice with OVA-induced asthma [35]. B-cells isolated from patients with allergy to birch pollen produce EVs presenting allergen peptides and thus activating specific T cells to proliferate and secrete Th2-associated cytokines, including IL-5 and IL-13 [57]. These EVs carry also leukotriene biosynthesis enzymes and enhance leukotriene and IL-8 production in HBEC [58]. Human DCs from cat-allergic blood donors can present the major cat allergen Fel d 1 to other immune cells and induce Th2-type cytokine production, including IL-4 [59]
Importantly, Bartel et al. determined also the levels of miRNAs in extracellular vesicles from nasal lavages of children with asthma. Lower expression of miR-34a, miR‐92b, and miR‐210 levels compared to healthy controls was connected with airways obstruction (Fig. 1). It was concluded that the above-mentioned miRNAs might play an important role in the dendritic cell maturation and development of the Th2 response in the early stage that leads to the asthma onset. Further complexity in epithelial EVs biology was demonstrated by Schindler et al., who analyzed EVs secreted by bronchial epithelial cells from healthy and asthmatic individuals [34]. Importantly, the biological activity of EVs is highly dependent on their cellular origin and route of secretion. Bronchial epithelial cells release distinct EV populations from apical and basolateral surfaces, which differ in size, molecular cargo, and functional properties (Fig. 3). Those secreted to the basolateral compartment were bigger, more abundant and had higher expression of CD9 and CD81 tetraspanin surface markers when compare to EVs released to the apical cell side. Analysis of miRNAs composition revealed in basolaterally secreted EVs the highest amounts of molecules from the miR-320, miR-181, miR-550, let-7, and the miR-154 families which were involved in the T and B cell receptors signalling pathways (Table 1). Contrary to that, miRNAs enriched in EVs from the apical side of the bronchial epithelial cells were connected with the mammalian target of rapamycin (mTOR) and mitogen-activated protein kinase (MAPK) signalling pathways [34]. This spatial heterogeneity clearly demonstrated the complexity of EV-mediated communication in asthma.
Environmental factors also influence EV-mediated mechanisms in asthma. Exposure to particulate matter (PM2.5) alters the miRNA cargo of epithelial-derived EVs, promoting pro-inflammatory signaling and airway dysfunction, thereby linking environmental factors with molecular mechanisms of asthma [35] (Table 1; Fig. 3).
Clinical studies have reinforced the relevance of EVs in asthma. Paredes et al. found that tetraspanins CD81 and CD63 were expressed in higher amounts in exosomes from BALF of birch pollen-allergic patients than in controls [58]. Owing to their enzymatic cargo involved in leukotriene biosynthesis, these exosomes enhanced leukotriene and IL-8 production in human bronchial epithelial cells, suggesting a direct contribution to asthma pathogenesis. Similarly, Levänen et al. [60] investigated miRNA profiles of exosomes from BALF obtained from asthmatic and healthy individuals. Their analysis revealed, that exosomes from asthmatic patients were enriched with 24 miRNAs compared to those isolated from controls, and these alterations were strongly correlated with lung function expressed as forced expiratory volume (FEV1) values. Furthermore, pathway analysis showed that the altered miRNA were linked to level of cytokines crucial for asthma pathogenesis such as IL-13, IL-10, IL-6, and IL-8, and were associated with the MAPK and JAK-STAT signalling pathways. Importantly, a subset of 16 miRNAs enabled a clear distinction between the asthmatic and healthy groups, suggesting that BALF-derived exosomes may represent a promising tool for asthma diagnostics [60].
In addition to noninvasive sources such as nasal lavage fluid and serum – often referred to as “liquid biopsies” – EVs involved in allergic processes can also originate from diverse immune cell populations. For example, dendritic cells stimulated with thymic stromal lymphopoietin (TSLP) were shown to release exosomes containing the OX40 ligand, a molecule that promotes Th2 differentiation, which represents a key process in asthma pathogenesis [61]. Similarly, neutrophils exposed to lipopolysaccharide (LPS) produce EVs enriched in neutrophil degranulation products, including proteases that promote myocyte proliferation. This process enhances airway smooth muscle proliferation, leading to asthma exacerbation [62]. Notably, eosinophil-derived EVs exert both autocrine and paracrine effects, amplifying inflammatory responses and impairing epithelial repair mechanisms [63]. Further evidence from animal models demonstrates that modulation of EV release influences disease severity [55]. The authors observed increased exosome secretion by bronchial epithelial cells and macrophages compared with other airway cell types (Fig. 2). Moreover, IL-13 modulated both the abundance and molecular composition of epithelial-derived EVs, thereby amplifying allergic inflammation through enhanced monocyte proliferation. Pharmacological inhibition of exosome release using GW4869 reduces airway inflammation and immune cell activation, although excessive inhibition may disrupt physiological homeostasis, indicating a dual role of EVs in maintaining immune balance [55]. As inhaled allergens first come into contact with alveolar macrophages and epithelial cells, their collaboration is crucial for appropriate inflammatory reactions and the preservation of homeostasis. Draijer et al. showed that alveolar macrophages deliver suppressor of cytokine signaling 3 (SOCS3) to alveolar epithelial cells via EVs, thereby contributing to the regulation of lung inflammation by inhibition of JAK-STAT signaling [64]. SOCS3 levels in bronchoalveolar lavage fluid were reduced in asthmatics and allergen-challenged mice. Alveolar macrophage–derived EVs containing SOCS3 inhibited STAT3/STAT6 activation and cytokine expression in epithelial cells, but this effect was lost under allergic inflammation. Administration of SOCS3 liposomes reduced lung inflammation in mice, suggesting SOCS3 delivery may represent a potential therapeutic strategy in asthma.
An equally significant aspect of exosomes’ functions in allergy is their potential role as carriers of aeroallergens. Admyre et al. demonstrated that B cell-derived exosomes possess antigen-presenting capabilities relevant to allergic responses [57] (Fig. 3). They showed that Epstein-Barr virus-transformed B cells isolated from patients with allergy to birch pollen released exosomes expressing high levels of MHC class I and II together with CD40, CD80, and CD86 costimulants, CD19 marker, as well as CD63 and CD81 tetraspanins. When loaded with peptides derived from the major birch pollen allergen Bet v 1, these exosomes induced dose-dependent proliferation of Bet v 1-specific T cells and promoted the secretion of Th2-associated cytokines, including IL-5 and IL-13. This study provided the first evidence that exosomes can present allergen-derived peptides and directly activate allergen-specific T cells, underscoring the contribution of APC-derived EVs to T cell-mediated allergic immune responses and highlighting their potential role in allergy pathophysiology. Vallhov et al. proved that exosomes derived from human dendritic cells (DCs) of cat-allergic blood donors could present the major cat allergen Fel d 1 to the other immune cells, stimulate Th2-like cytokine production (IL-4) in allergic donors [59] (Fig. 3). This finding may have practical implications for the development of novel immunotherapeutic and exosome-based vaccines. Further insights into eosinophil-derived EVs were provided by Canas et al., who compared exosomes isolated from blood eosinophils of asthmatic patients and healthy donors [53]. Exosomes from asthmatic individuals increased ROS and NO production, enhanced eosinophil adhesion to fibronectin, and promoted cell viability, indicating a potential autocrine mechanism that amplifies eosinophil-driven inflammation. Subsequent studies by the same group revealed that these exosomes also induced apoptosis in human small airway epithelial cells and delayed epithelial wound repair [54]. These effects were associated with altered PI3K/AKT and JAK-STAT signaling, as evidenced by reduced phosphorylation of AKT and STAT3 (Fig. 1). In addition, eosinophil-derived exosomes increased the expression of proinflammatory genes, including TNF, CCL26, and POSTN, and promoted bronchial smooth muscle cell proliferation via enhanced MAPK signaling. Collectively, these findings demonstrate the pleiotropic role of eosinophil-derived exosomes in asthma pathogenesis, airway remodeling, and disease exacerbation [54]. Lv et al. used papain-induced asthmatic mouse model to investigate the possible EVs-mediated interaction between the most important populations of innate immune cells in airway tissues specifically, lung M2 macrophages and group 2 innate lymphoid cells (ILC2s) [65]. Immunofluorescence and flow cytometry analyses revealed, that after intratracheal injection of lung M2 macrophage-derived EVs to asthmatic mice, ILC2s were capable of recognizing and interacting with these vesicles. Furthermore, ILC2s can use various endocytic pathways for EVs uptake, allowing them to adapt to changing environmental conditions. Additionally, separated mouse CD4+ T cells and macrophages cultured with PKH26-labeled M2 EVs for 24 h also internalized the vesicles, suggesting their indirect ability to promote the activation of ILC2s through macrophages and CD4+ T cells. RNA sequencing analysis identified the transcript 4930474H06RiK as a potential mediator of ILC2s activation in allergic airway inflammation, as its expression was highly increased in M2 macrophage-derived EVs from the lung tissue of and asthmatic mice compared with control animals. Functional analyses indicated that this transcript promotes cytokine production and ILC2 proliferation, possibly through downregulation of glycolysis-related molecules such as HK2. These results highlight the role of M2 macrophage-derived EVs in amplifying type 2 immune responses in asthma.
Interestingly, EVs of non-host origin, such as those derived from dietary sources, may also modulate immune responses. For example, bovine milk-derived EVs have been shown to influence macrophage polarization and alter inflammatory responses in the lung, suggesting that exogenous EVs may contribute to the regulation of airway inflammation [56]. Macrophages isolated from mice fed intact milk EVs and stimulated ex vivo with aques extracts of agricultural dust produced higher levels of M1-associated proinflammatory cytokines, including tumor necrosis factor alpha (TNF-α), IL-12/23, and IL-6, while the expression of M2-associated anti-inflammatory mediators such as IL-10 and arginase was reduced (Fig. 2). To sum up, findings demonstrate that EVs are central regulators of asthma pathogenesis, acting through multiple mechanisms including miRNA transfer, immune cell modulation, antigen presentation, and environmental signal integration. Their effects are highly context-dependent, influenced by cellular origin, microenvironment, and disease stage, which underscores both the complexity and therapeutic potential of EV-targeted strategies in asthma.
Extracellular Vesicles in the Development of Atopic DermatitisAtopic dermatitis (AD) is a chronic inflammatory skin disease characterized by barrier disruption, immune dysregulation, and recurrent eczema accompanied by intense pruritus and xerosis. It is one of the most common inflammatory disorders in childhood, affecting up to 20% of the pediatric population, and often precedes the development of other atopic diseases, including asthma, allergic rhinitis, and food allergy. Increasing evidence also suggests that AD has systemic consequences extending beyond the skin [66].
The epidermis is central to AD pathogenesis. Under physiological conditions, its stratified structure and tightly regulated keratinocyte differentiation ensure effective protection against water loss, allergens, irritants, and microorganisms. In AD, however, impaired barrier integrity, abnormal keratinocyte differentiation, and altered immune signaling facilitate allergen penetration and microbial colonization. This is accompanied by a type 2-skewed inflammatory milieu, characterized by increased expression of IL-4, IL-5, IL-13, IL-22, and IL-31, elevated IgE production, eosinophilia, and progressive tissue damage [67].
The regulation of the skin microenvironment relies on a complex interplay between blood, skin cells, immune cells, skin microbiome. Within this context, EVs have emerged as important mediators of this communication. In AD, EVs appear to contribute to disease development through three major mechanisms: microbial induction of skin inflammation, modulation of keratinocyte homeostasis, and regulation of antigen-specific immune responses.
Commensal and pathogenic microorganisms residing on the skin surface are capable of releasing EVs that deliver bioactive cargo to host cells, thereby modulating local immune responses. One of the best-characterized examples involves Staphylococcus aureus-derived EVs [68]. In a murine model, topical application of gauze soaked with S. aureus EVs onto barrier-disrupted skin induced epidermal thickening and dermal infiltration by mast cells and eosinophils. Furthermore, T cells isolated from skin-draining lymph nodes exhibited increased production of IFN-γ and IL-17 (Fig. 4).
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Contribution of Staphylococcus aureus–derived EVs to atopic dermatitis pathogenesis. Bacteria that constitute the skin microbiota can release extracellular vesicles (EVs) capable of delivering a variety of bioactive molecules to host cells, thereby initiating inflammatory responses. In a murine model, application of gauze soaked with Staphylococcus aureus–derived EVs to tape-stripped skin resulted in epidermal thickening and increased dermal infiltration of mast cells and eosinophils. Additionally, T cells isolated from skin-draining lymph nodes of EV-treated mice exhibited elevated production of IFN-γ and IL-17. In vitro stimulation of primary mouse dermal fibroblasts with S. aureus EVs led to enhanced secretion of proinflammatory mediators, including IL-6, TSLP, macrophage inflammatory protein-1α (MIP-1α), and eotaxin [68]. Furthermore, S. aureus EVs were shown to contain the virulence factor α-hemolysin, which promotes keratinocyte necrosis by upregulating IL-6 production [
Comments (0)