
Remember me
There have been around 300 patients reported with IL-10 signalling since the first description in 2009 [9]. Based on the large cohort study and a systematic review, most patients have IL10RA defects, followed by mutation in IL10RB and IL10 [9, 10]. The most common variants in IL10RA are homozygous missense mutation or heterozygous missense mutations. Among all mutations, the IL10RA c.C301T (p.R101W and c.G537A (p.T179T) mutations are the most common mutations [10]. Both mutations are significantly more prevalent among patients from East Asia than among those of other regions [10]. There is no significant difference in disease onset time, perianal lesion involvement, or mortality rate among patients with IL10, IL10RA or IL10RB deficiencies, however, B-cell lymphoma is more commonly associated with IL10RB deficiency [9, 10].
When proband clinical exome sequencing only identified one heterozygous pathogenic variant in the IL10RA among a VEOIBD patients, trio genome sequencing showed an additional novel deletion spanning exon 1 (c.-149_67 + 117del) [11]. Subsequently, another five patients were found to have this 333 bp deletion spanning exon 1 of the IL10RA gene and another missense mutation, and one patient has homozygous mutation of the 333 bp deletion in IL10RA [11]. Later another study also reported four VEOIBD patients with the same deletion [12]. Because the 333 bp deletion cannot be captured by the routine analysis of clinical exome sequencing or whole exome sequencing, these patients had delayed genetic diagnosis. More recently, Shi et al. reported the first case of apparent homozygous mutations in IL10RA [13]. Using a combination of whole-exome sequencing, comparative genomic hybridization and SNP array, a paternal uniparental diploidy (c.301 C > T/11q12.3-11q25del) [upd (11) pat] on chromosome 11 was identified in a VEOIBD patient. Together with earlier studies, it is essential to apply additional sequencing methods such as genome sequencing when initial genetic analysis is inconclusive, as exon-based approaches are limited to detect deletions or duplications. Notably, large deletions have also been identified in IL10RB, while no phenotypic differences are shown in patients with large deletions and other types of mutations [9].
There is also a geographic accumulation of confirmed pathogenic IL10RA variants in East Asia [14]. Possible explanation of high frequency of a rare genetic disease in a certain population includes founder effect, genetic drift or selective advantage [15]. Based on the geospatial association analysis between pathogenic IL10RA variants and human pathogen distribution worldwide, Schistosomiasis japonicum might be the cause of long-term pathogen-mediated natural selection pressure for these variants in East Asia [14]. Interestingly, a recent study identified a founder effect of IL10RB W40X mutation using low-pass whole genome sequencing data and haplotype analysis surrounding IL10RB in several members of the pedigree from an indigenous group living in a remote region of Ecuador [16]. This is consistent with another study, which reported a copy number variation with founder effect in IL10RB in the Portuguese kindreds [17]. Collectively, these results highlight the necessity of advanced sequencing strategies to provide accurate genetic diagnosis and to understand the population genetics and the evolutionary consequences of these mutations.
Pathway and MechanismDefective Anti-Inflammatory Function in IL-10 Signalling DefectsWhen IL-10 binds to the its receptors, IL-10RA and IL-10RB, Janus tyrosine kinases, JAK1 and Tyk2 are activated, resulting in the phosphorylation of STAT3 (signal transducer and activator of transcription 3) and the induction of STAT3-dependent genes, including SOCS-3 [18]. Expression of SOCS-3 inhibits various inflammatory cytokines including TNF, IL-6, and IL-1β [19]. In the context of IL-10 signalling defects, peripheral blood mononuclear cells (PBMCs) from patients could not induce IL-10-mediated STAT3 phosphorylation or inhibition of TNF-induced proinflammatory cytokine secretion [4]. The abrogated IL-10 signaling leads to hyperinflammation, which is consistent with the extensive intestinal inflammation, severe perianal diseases presented in patients with IL-10 signalling defects [4, 20].
Phenocopy of IL-10 Signalling Defects Due to Anti-IL-10 AutoantibodiesHigh titre neutralizing autoantibodies against IL-10 was identified in infantile-onset IBD patients without pathogenic mutations detected on genetic sequencing [21]. Functional assay showed unresponsiveness of patient’s PBMCs to IL-10, while normal response was confirmed upon isolation by density gradient centrifugation and thorough washing of the PBMCs. Therefore, phenocopy of the IL-10 signalling defects were suggested due to the high titer of auto-antibodies to IL-10 [21]. One patient had disease onset at 3 months with severe IBD phenotype and another patient presented with moderate pancolitis from 4.5 years of age. Interestingly, both patients had high titre of anti-IL-10 autoantibodies which blocked the IL-10-dependent downregulation of the TNF-α response to LPS [21]. B cell-depleting therapy with rituximab combined with intravenous immunoglobulin led to clinical response in one patient, while the other patient with milder course did not require B cell-depleting therapy [21]. Previously, higher levels of anti-IL-10 antibodies were found in 17% of the adult patients with CD [22]. Together with earlier findings, further studies are required to investigate the role of anti-IL10 autoantibodies in paediatric and adult IBD, which phenocopy the monogenic IBD caused by IL-10 signalling defects. These studies show that pathogenesis observed in the rare monogenic diseases might be generalisable to the non-genetic diseases which affect a larger populations and provide further implications for developing targeted therapy in selected patients. The comparisons of charateristics of patients with IL-10 signalling defects and IBD patients with anti-IL-10 autoantibodies are shown in Table 1.
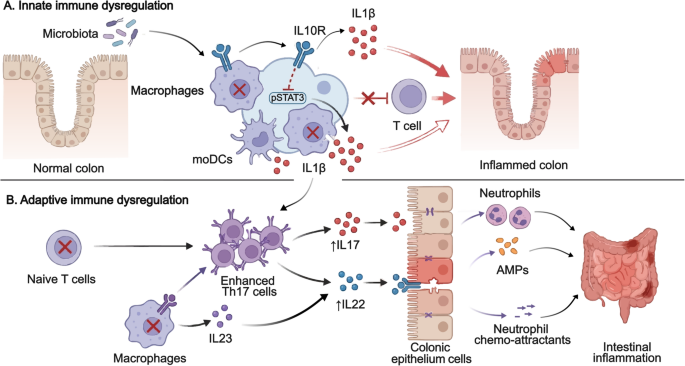
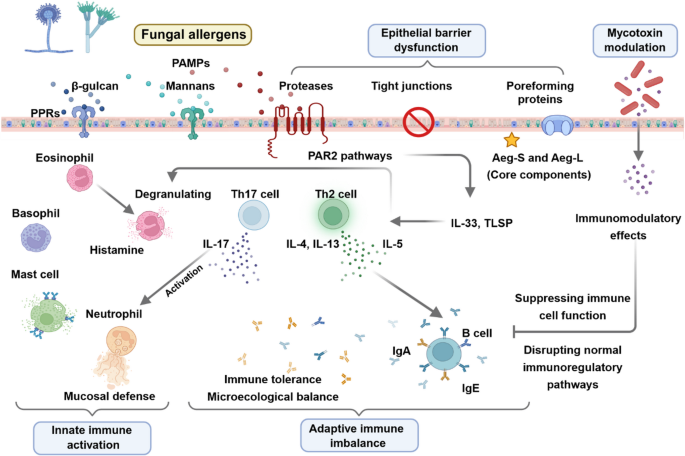
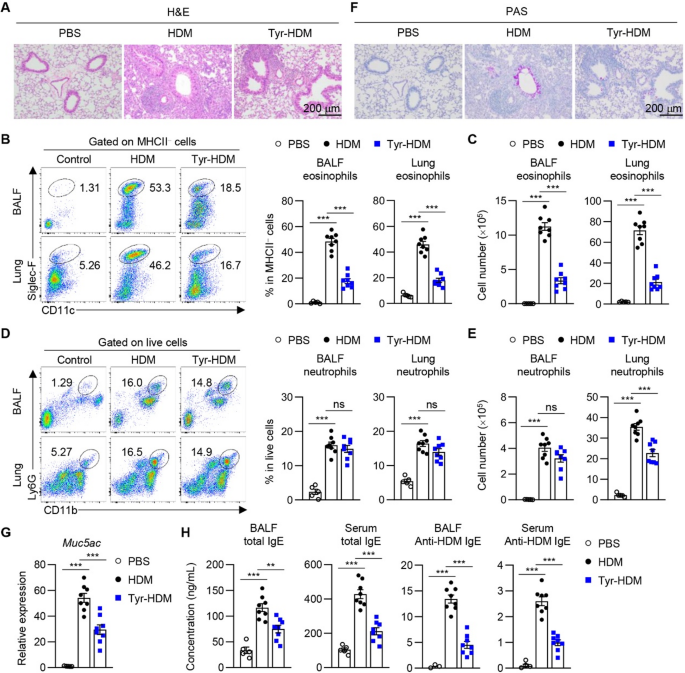
Table 1 Comparison of characteristics of patients with IL-10 signalling defects and IBD patients with anti-IL-10 autoantibodiesInnate Immune Dysregulation in IL-10 Signalling DefectsBoth innate and adaptive immune dysregulation have been observed in IL-10 signalling defects (Fig. 1). Cytokines produced by innate and adaptive immune cells contribute to the pathogenesis of IBD [23]. Increased inflammatory cytokine IL-1β is found in active IBD and IL-1β is involved in the colitis development and Th17 associated response in the intestine [24]. Stimulation of murine and human IL10R-deficient macrophages by LPS + ATP led to increased IL-1β production which was not suppressed by IL-10 pre-treatment [25]. Furthermore, LPS stimulation alone increased IL-1β secretion via noncanonical, caspase 8-dependent activation of the inflammasome in the human IL10R-deficient macrophages [25]. Notably, there is signiciant differences in innate immune regulation between the human and murine IL10R-deficient macrophages. Macrophages derived from patients are capable of producing IL-1β upon LPS activation without the secondary inflammasome activatin trigger (for example ATP) [25]. Overall, this finding suggested that blocking IL-1 in patients with IL-10 signalling defects may be beneficial, which has been supported by successful treatment outcomes with anakinra in two patients. Cellular metabolism in macrophages have shown profound changes in the metabolic profiles during macrophage activation and it is essential for regulating inflammation and tissue repair [26]. Macrophages accumulate damaged mitochondria in Il10–/– mice and IL10R-deficient patients, which resulted in dysregulated activation of the NLRP3 inflammasome and production of IL-1β [27]. Inhibition of ROS or mTOR signalling by antioxidants or rapamycin, can suppress the IL-1β secretion in the monocyte-derived macrophages from IL10R-deficient patients [27]. Consequently, targeting the mTORC1 pathway in macrophages could be beneficial for treatment or prevention of IBD. Interestingly, time-series experiments using Il10rb−/− mice showed that intestinal inflammation and macrophage dysfunction started during the third week of life, coinciding with weaning and the associated diversification of the intestinal microbiota [28]. As a large cohort study on patient with IL-10 signalling defects showed that 70.3% (97 of 138) had onset of disease during neonatal period and only 2.2% (3 of 138) had onset after 1 year old, understanding the timing and development of colitis using murine model would be essential [10]. Taken together, these results indicate that IL-10R-mediated regulation of macrophage function during the early postnatal period might be the critical time window leading to the development colitis.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Innate and adaptive immune dysregulation in IL-10 signalling defects. Both innate and adaptive immune dysregulation are associated with the pathogenesis of IL-10 signalling defects. The defective macrophages and monocyte-derived dendritic cells (moDC) produce increased IL1β, which lead to intestinal inflammation. IL10R-deficient T naive cells exhibit increased proliferation and enhanced Th17 response. The defective macrophages produce IL23, which recruits TH17 cells and induces secretion of IL22 to cause intestinal inflammation. AMP: antimicrobial peptide, moDC: monocyte-derived dendritic cells.
Another study revealed that IL-10 signalling in monocyte-derived dendritic cells (moDCs) is crucial for controlling IFNγ-secreting CD4 + T cells [29]. IL-10 signalling defects significantly increased IL-1β release by moDCs [29]. Subsequently, a subgroup of paediatric IBD patients having higher IL-1β expression in activated immune cells and affected intestinal tissue were identified, together with a subgroup of patients exhibited diminished IL-10 responsiveness [29]. Notably, this finding corroborates with another study, for which IL-10R expression and IL10-induced pSTAT3 were decreased in monocytes from patients with Crohn’s Disease (CD) [30]. Collectively, these results highlighted the important role of IL-10 signalling in the moDCs and monocytes and IL-1β as a potential classifier for IBD patients.
Adaptive Immune Dysregulation in IL-10 Signalling DefectsIL10R-dependent signals have been proved to be critical for the regulatory and effector CD4 + T-cell function in mice [31]. IL10R-deficient T naive cells exhibit increased proliferation and enhanced Th17 response [32]. Increased innate immune production of IL-1β in IL10R deficiency leads to enhanced production of IL-17 A [32]. Concurrently, IL10RA-deficient gut macrophages produce IL-23, which recruits pathogenic Th17 cells and induces secretion of IL-22 in the Cx3cr1cre:Il10rafl/fl mice model [33, 34]. Subsequently, colonic ECs respond to IL-22 exposure by expression of antimicrobial peptide (AMPs) and neutrophil chemo-attractants that drive a proinflammatory epithelial cell response model [34]. Although IL-22 is generally considered to be protective for intestinal inflammation and might be involved in wound healing [35]. The protection from colitis observed in IL22-deficient Cx3cr1cre:Il10rafl/fl mice indicates the proinflammatory role of IL-22 in the context of IL-10 signalling defects. In contrast, anakinra treatment led to mucosal healing associated with increased frequency of IL-22-producing lymphocytes in the lamina propria of the terminal ileum in a patient with IL10RA mutations [36].
Beyond dysregulated cytokine production, impaired IL-10 signalling also affects the adaptive immune compartment at the level of antigen receptor diversity. Next generation sequencing of the T and B cell receptor repertories demonstrates the adaptive immune function in many immune-mediated disorders, including UC, juvenile idiopathic arthritis and Wiskott-Aldrich syndrome [37,38,39]. Both the T and B cell receptor repertoires were skewed in patients with IL-10 signalling defects and are characterized by enhanced clonality, and alterations in repertoire features especially in T cells [40]. These findings are consistent with the studies of IBD without underlying genetic defects and other chronic intestinal inflammation [41, 42].
Collectively, these results provide mechanistic insights into the adaptive immune dysregulation triggered by the IL-10 signalling defects. Targeting the key cytokines may be beneficial for suppressing intestinal inflammation and inducing mucosal healing in such patients.
Microbiota and in IL-10 Signalling DefectsThere is a significant association between gut microbiota and IBD, for example, relative increase in Proteobacteria, mainly E. coli, was identified in CD patients [43]. Environmental factors might modify the variable expressivity, progression, severity, and onset of some monogenic diseases, including Cystic fibrosis, Huntington disease [44]. Interestingly, germ free IL10-deficient mice did not develop colitis, which suggested that normal enteric bacteria are essential for the development of spontaneous colitis in IL-10 signalling defects [45]. .
The changes in diversity and composition in the gut microbiota of IL10-deficient mice during the onset and progression of colitis were initially described. Increase in Proteobacteria and Escherichia coli were reported in IL10-deficient mice [46]. Subsequently, the relative abundance of Firmicutes was significantly increased in patients with IL10RA defects compared to healthy controls [47]. Another study showed a significant decrease in the abundance and diversity of the gut microbiota among IL10RA-deficient patients compared to those with late-onset paediatric IBD and non-monogenic VEOIBD [48]. The author also identified genus Bifidobacterium as a potential diagnostic indicator for different groups of IBD [48]. Furthermore, association between microbiota and engraftment failure has also been examined in patients with IL10RA defects undergoing umbilical cord blood transplantation (UCBT). Patients achieved successful engraftment had a higher level of Lautropia genus that sustained throughout the UCBT and had significantly higher Shannon diversity values on the UCBT day [49].
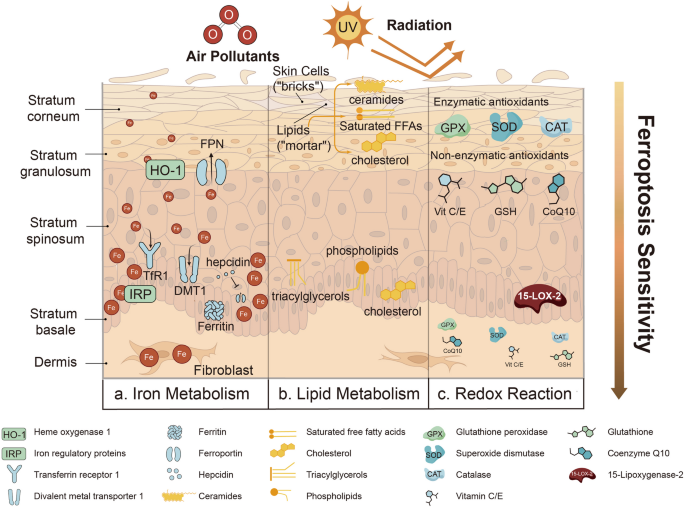
Therapeutic StrategiesA wide range of classic IBD treatment have been tested in patients with IL-10 signalling defects. As the advanced understanding of the underlying pathogenesis, more targeted therapy has been considered (Fig. 2).
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Current and prospective treatment options for the IL-10 signalling defects. Current treatment options include immunomodulator, biologics, surgery and haematopoietic stem cell transplant. Thalidomide and interleukin 1 receptor antagonist might lead to partial response. Surgical intervention especially enterostomy is a bridging therapy which can significantly improve quality of life. HSCT can be curative to the patients and allows subsequent stoma closure. Gene therapy is only available in the murine model and further clinical trials are required. HSCT: haematopoietic stem cell transplant, IL1R: interleukin 1 receptor, IL10: interleukin 10, UCBT: umbilical cord blood transplant.
ImmunomodulatorClassic medical therapies suppress the immune system, which helps control intestinal inflammation [50]. Many immunomodulators targeting various pathways, including methotrexate [20], azathioprine [51], ciclosporin [51], tacrolimus [20] and mercaptopurine [52] have been tried in patients with IL-10 signalling defects, but almost all have been proved to be ineffective. Thalidomide might be an alternative option for refractory paediatric IBD with significant remission and clinical improvement based on a narrative review of 14 studies involving 213 children including 16 cases with IL-10 signalling defects [53]. Thalidomide has been reported in several studies and most patients with IL-10 signalling defects showed partial response to the treatment [20, 54,55,56]. A recent retrospective cohort study showed that thalidomide is effective and tolerable in children with VEOIBD, however, monogenic patients were excluded [57]. However, thalidomide is associated with multiple adverse effects including peripheral neuropathy, sedation and constipation [58], which have been reported in patients with IL-10 signalling defects [51,
Comments (0)