
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Remember me
Oxidative stress increases with aging and may influence both sleep physiology and gut microbial activity. Molecular hydrogen, produced by intestinal fermentation, acts as an endogenous antioxidant and can be measured noninvasively in exhaled breath. However, age- and sex-related differences in nocturnal changes of breath hydrogen remain unclear. We analyzed breath hydrogen and methane levels in 166 healthy adults aged 20–85 years. Participants self-collected end-alveolar breath samples at home before sleep and immediately after waking. Breath hydrogen and methane levels were determined by gas chromatography. The older group (n = 91) showed significantly lower breath hydrogen levels after waking compared with the non-older group (n = 75), despite no differences before sleep. No significant sex-related differences were observed. The older group also reported more frequent nocturnal awakenings. These findings suggest that age-related changes in hydrogen dynamics may be associated with sleep-related physiological processes. The overnight change in breath hydrogen was significantly greater in the older group compared with the non-older group, indicating a larger overnight reduction in hydrogen levels in older adults. No significant differences in breath methane levels were observed between groups. Age-related alterations in gut microbiota, gastrointestinal motility, and redox balance may contribute to reduced morning hydrogen levels. Breath hydrogen measurement represents a simple, noninvasive biomarker for assessing physiological changes associated with aging and sleep.
Export citation and abstractBibTeXRIS
Oxidative stress, caused by an imbalance between reactive oxygen species production and the body’s antioxidant defense, plays a key role in aging, cardiovascular diseases, and cognitive decline. Identifying endogenous factors that reduce oxidative stress is crucial for understanding these conditions. Among such potential factors, Neale. First proposed that molecular hydrogen, produced during intestinal anaerobic fermentation, has reductive properties [1]. This hypothesis provided an important perspective, suggesting that a naturally occurring molecule may influence oxidative balance, yet it attracted limited attention in subsequent clinical research.
In 2007, Ohsawa et al demonstrated that molecular hydrogen selectively reduces hydroxyl radicals and alleviates ischemia-reperfusion injury, representing an important advance in hydrogen research [2]. Since then, an expanding body of literature has reported on the efficacy of hydrogen therapy in various clinical applications, underscoring its therapeutic potential across multiple organ systems [3, 4]. Beyond modulating oxidative stress, molecular hydrogen exerts preventive and therapeutic effects, partly through its antioxidative actions such as neutralizing hydroxyl radicals and reducing peroxynitrite levels [5].
Hydrogen gas detected in exhaled breath originates mainly from the anaerobic fermentation of residual dietary components, particularly fiber-rich substrates, by the gut microbiota [6]. In the colon beyond the ileocecal valve, anaerobic bacteria ferment these undigested residues to produce molecular hydrogen, which is rapidly absorbed into the bloodstream and subsequently excreted via the lungs, allowing its measurement in exhaled breath. The area under the curve (AUC) of breath hydrogen level over time reflects the amount of hydrogen produced through intestinal anaerobic fermentation [7]. This AUC is proportional to the total extent of dietary fiber degradation by colonic microbiota, establishing it as a valuable marker of fermentation activity [8].
The gut microbiota, composed of diverse bacterial populations in the colon, is strongly influenced by age [9], as well as by factors such as diet, psychological stress [10], and physical activity [11]. With advancing age, the diversity and metabolic activity of the gut microbiota undergo significant changes, including reduced microbial diversity and altered metabolic profiles, which can affect gastrointestinal function and systemic health. These age-related alterations are increasingly recognized as contributors to a wide range of conditions, including obesity, metabolic disorders, mood disturbances through brain-gut interactions [12], and cardiovascular diseases [13].
Several studies have reported an association between aging and the level of hydrogen gas in exhaled breath. For example, a study of Japanese centenarians found that their exhaled hydrogen levels were higher than those of comparatively non-older healthy individuals [14]. This observation may be explained by an age-related increase in intestinal hydrogen production, which contributes to the elevated breath hydrogen levels observed in older adults [15]. However, because oxidative stress is also elevated with aging [16], hydrogen utilization as an antioxidant may increase [5], suggesting a dynamic balance between its production and consumption.
Sleep duration and quality decline with advancing age, with older adults often experiencing longer sleep onset latency, increased fragmentation, and reduced efficiency [17, 18]. Traditionally, sleep quality has been assessed using self-reported questionnaires such as the Pittsburgh sleep quality inventory [19]. However, such subjective assessments are prone to recall bias and may not accurately capture physiological sleep states. To overcome these limitations, objective methods, including polysomnography, heart rate variability, and actigraphy, have been increasingly employed to evaluate sleep quality through physiological signals [20–22]. These non-invasive and cost-effective approaches offer more objective and reliable insights into age-related changes in sleep.
Recent studies have highlighted the relationship between gut microbiota and circadian rhythms. The composition and metabolic activity of the gut microbiome exhibit diurnal variations, and microbial metabolites have been shown to influence clock gene expression in both the central nervous system and the liver [23, 24]. Through these mechanisms, the gut microbiota may contribute to the regulation of sleep-wake cycles and energy balance. Among these microbial metabolites, hydrogen gas is a notable byproduct of intestinal fermentation that reflects gut function and digestion. Given its physiological relevance, exhaled hydrogen may serve as a non-invasive indicator of microbiota-related changes associated with sleep.
In this study, we aimed to investigate whether breath analysis could serve as a complementary, non-invasive approach to assessing sleep quality. Although breath hydrogen measurement is non-invasive and practical, baseline values before and after sleep, particularly across different age groups, have not been established. To address this gap, we measured breath hydrogen levels in older and non-older adults and compared differences between the groups. The aim of this study was to investigate age-related differences in overnight changes in breath hydrogen and to explore their potential physiological relevance.
2.1. ParticipantsA total of 166 community-dwelling adults aged 20–85 years were recruited through local advertisements. Participants were divided into a non-older group (20–59 years, n = 75) and an older group (60–85 years, n = 91). Individuals taking α-glucosidase inhibitors were excluded because these medications are known to increase breath hydrogen levels by enhancing intestinal fermentation. Participants with known gastrointestinal disorders or other conditions judged by the investigators to interfere with breath gas measurements were also excluded.
2.2. ProtocolsOn the day before the study, participants visited the laboratory and received verbal and written instructions on breath sample collection. They were asked to refrain from alcohol consumption before the measurements and to maintain their usual daily routine. At home, participants collected breath samples immediately before going to sleep and immediately after waking in the morning. They also completed a sleep diary, recording bedtime, wake-up time, sleep duration, and the number of nocturnal awakenings. The collected samples were returned to the laboratory for analysis [22, 25].
2.3. Breath hydrogen sampling and analysisEnd-alveolar breath was collected in a 400 ml aluminum-pouched sampling bag (Collection Bag, Laboratory for Expiration Biochemistry, Nourishment Metabolism Co., Ltd, Nara, Japan). Before the study, all participants underwent training in the breath collection procedure during a laboratory visit. Participants were instructed to inhale normally, hold their breath for approximately 15 s, and then exhale fully into the sampling bag to obtain end-alveolar breath. To further minimize contamination from dead-space air, participants were instructed to exhale completely after breath-holding.
Breath samples were collected at home immediately before sleep and immediately after waking and were returned to the laboratory in sealed sampling bags. All samples were analyzed within approximately 10–12 h of collection. Immediately before analysis in the laboratory, each breath sample was transferred into a gas-tight glass syringe, and 1 ml of the sample was injected into a gas chromatograph equipped with a semiconductor detector (TRIlyzer mBA-300; Taiyo Ltd, Osaka, Japan) to measure breath hydrogen and methane. Hydrogen and methane were separated from other compounds through a column packed with molecular sieve and detected by an SnO2 semiconductor sensor [26].
Ambient air samples were collected in separate sampling bags of the same type for each measurement session and analyzed in the same manner. Breath hydrogen and methane levels were calculated by subtracting the corresponding ambient air values.
Previous studies have shown that breath hydrogen remains relatively stable under similar storage conditions, although some degree of variability due to gas diffusion cannot be completely excluded [27–29].
2.4. Statistical analysesΔH2 and ΔCH4 were calculated as values before sleep minus those after waking. Group comparisons were performed using Student’s t-test or the Mann–Whitney U test, as appropriate. Associations between ΔH2 and demographic variables (age and sex) and gastrointestinal symptom scores from the digestive tract section of the Cornell Medical Index (CMI) were examined using Spearman’s correlation or the Mann–Whitney U test. Variables that showed significant associations in univariate analyses were entered into a multivariate logistic regression model, with positive or negative ΔH2 as the dependent variable. Methane levels were analyzed in the same manner as hydrogen levels for comparisons by age group and sex. Data are presented as mean ± standard deviation (SD). Effect sizes were calculated using Cohen’s d. Statistical significance was set at p < 0.05. All analyses were performed using SPSS version 31.0 (IBM Corp., Armonk, NY, USA).
3.1. Participant characteristicsThe non-older group (n = 75; mean age 43.1 ± 12.3 years; 21 men, 54 women) and the older group (n = 91; mean age 71.0 ± 5.9 years; 38 men, 53 women) did not differ significantly in body mass index or total sleep duration (table 1). However, the older group reported significantly more frequent nocturnal awakenings compared with the non-older group (p < 0.05).
Table 1. Participant characteristics in the non-older and older groups.
VariableNon-older group (n = 75)Older group (n = 91)p-valueMean age (years)43.1 ± 12.371.0 ± 5.9—Sex (me/women)21/5438/53—Sleeping hours (hours)7.1 ± 1.57.3 ± 1.5n.s.Body mass index (kg m−2)22.0 ± 2.822.8 ± 2.6n.s.Nocturnal awakenings (frequency)0.5 ± 0.71.1 ± 0.7p < 0.05Data are presented as mean ± SD. No significant differences were observed in body mass index or sleep duration, whereas the older group reported significantly more nocturnal awakenings than the non-older group (p < 0.05).
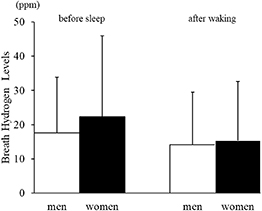
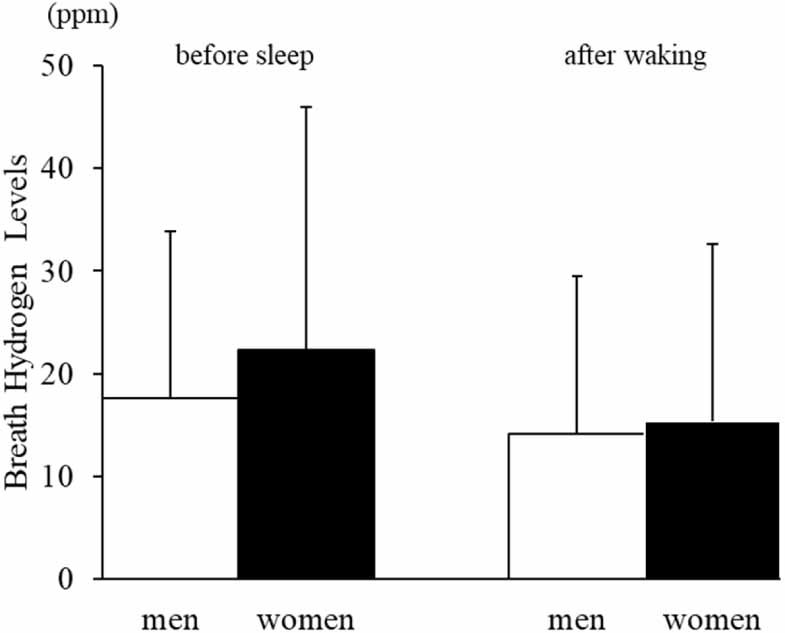
3.2. Breath hydrogen and methane levelsNo significant sex-related differences were observed in breath hydrogen levels either before sleep (men: 17.6 ± 21.0 ppm; women: 22.4 ± 19.7 ppm) or after waking (men: 14.1 ± 19.5 ppm; women: 15.3 ± 14.9 ppm; figure 1).
Figure 1. Breath hydrogen levels before sleep and after waking in men and women.Mean ± SD values for breath hydrogen levels are shown for men (white bars) and women (black bars). Although women tended to exhibit slightly higher values before sleep, no statistically significant sex-related differences were observed at either time point.
Download figure:
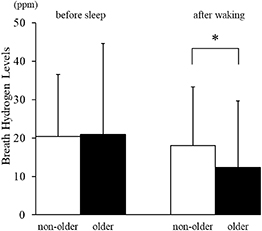
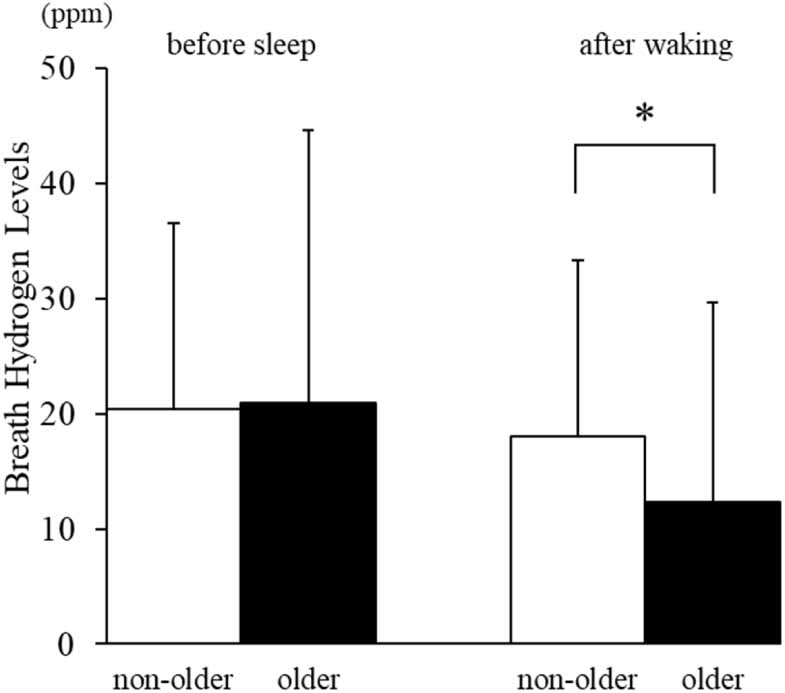
Standard image High-resolution imageWhen comparing age groups, no significant difference was observed in breath hydrogen levels before sleep (non-older: 20.4 ± 16.2 ppm; older: 20.9 ± 23.6 ppm). However, after waking, the older group (12.3 ± 17.3 ppm) exhibited significantly lower breath hydrogen levels than the non-older group (18.0 ± 15.3 ppm; p < 0.01, Cohen’s d = 0.37; figure 2).
Figure 2. Breath hydrogen levels before sleep and after waking in non-older and older adults.Mean ± SD breath hydrogen values are shown for the non-older (white) and older (black) groups. No significant differences were observed before sleep; however, older adults exhibited significantly lower breath hydrogen levels after waking compared with non-older adults (p < 0.01, Cohen’s d = 0.37).
Download figure:
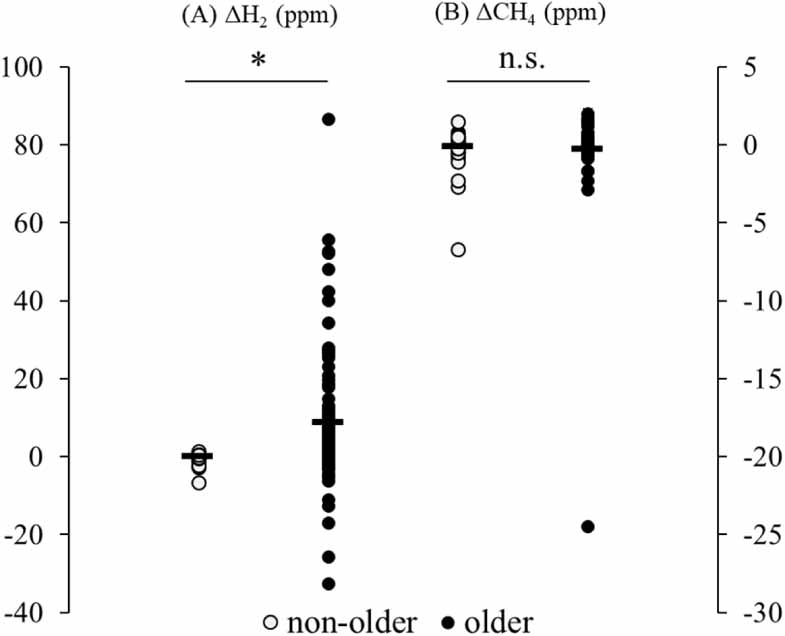
Standard image High-resolution imageΔH2 was significantly greater in the older group compared with the non-older group (p < 0.01, Cohen’s d = 0.35; figure 3(A)).
Figure 3. Changes in breath hydrogen (ΔH2) and methane (ΔCH4) levels between before sleep and after waking.(A) ΔH2 was significantly greater in older adults compared with non-older adults (p < 0.01, Cohen’s d = 0.35).(B) No significant differences were observed in ΔCH4 between groups.Each dot represents an individual participant. Horizontal bars indicate mean values.
Download figure:
Standard image High-resolution imageBreath methane levels did not differ significantly between groups either before sleep (non-older: 2.31 ± 1.28 ppm; older: 3.00 ± 3.15 ppm; p = 0.42) or after waking (2.43 ± 2.03 ppm vs. 3.29 ± 4.64 ppm; p = 0.26). ΔCH4 also did not differ significantly between the groups (figure 3(b)).
3.3. Correlation analysisIn univariate analyses, ΔH2 showed a significant negative correlation with age (p < 0.01) and with gastrointestinal symptom scores from the digestive tract section of the CMI (p = 0.04). However, this association was not retained in multivariate analysis.
The main finding of this study was that older adults exhibited significantly lower breath hydrogen levels after waking compared with non-older adults, despite no evident difference immediately before sleep. This pattern indicates that age-related differences are more pronounced in overnight hydrogen dynamics rather than in baseline levels, suggesting that physiological processes occurring during sleep contribute to the observed reduction in morning hydrogen.
Breath hydrogen reflects the net balance between intestinal production and multiple hydrogen-consuming pathways within the gut ecosystem [6–8]. Hydrogen is generated through microbial fermentation of unabsorbed carbohydrates, while being simultaneously consumed by processes such as methanogenesis and other hydrogenotrophic pathways [30, 31]. Therefore, changes in breath hydrogen should be interpreted as shifts in this dynamic balance rather than as a simple decrease in production alone.
Methane production represents an important alternative explanation for reduced breath hydrogen, as methanogenic archaea utilize hydrogen to produce methane [6]. In addition, the prevalence of methane producers has been reported to increase with age [32], making this mechanism particularly relevant in older populations. In the present study, methane was measured using the same breath samples; however, no significant differences were observed between age groups or sexes, and ΔCH4 did not differ between groups. These findings suggest that methane production is unlikely to fully explain the observed age-related reduction in overnight hydrogen. Nevertheless, we cannot exclude the possibility of temporal variation in methanogenesis or individual differences that were not captured by the two sampling time points.
Age-related changes in gut microbiota and gastrointestinal function may also contribute to the observed findings. Aging is associated with reduced microbial diversity and altered metabolic activity [9], which may influence hydrogen production. In addition, decreased gastrointestinal motility with aging may affect gas production, absorption, and excretion during sleep [33]. Taken together, these factors suggest that the lower morning hydrogen levels observed in older adults may reflect an age-related shift in the balance between hydrogen production and consumption.
Sleep-related physiological changes provide another plausible explanation. Gut microbial activity exhibits diurnal variation [23, 24], and sleep is associated with coordinated changes in autonomic nervous system activity, gastrointestinal motility, and metabolic regulation [34]. Older adults often experience fragmented sleep and altered sleep architecture [17, 18], which may disrupt normal nocturnal gastrointestinal function. In the present study, older adults also reported more frequent nocturnal awakenings, which may have contributed to altered gas handling and reduced morning hydrogen levels.
Dietary factors, particularly fiber intake, are known to influence breath hydrogen levels and contribute to interindividual variability [6, 8]. However, dietary intake was not controlled in this study. According to the National Health and Nutrition Survey in Japan (2023), average dietary fiber intake is relatively comparable between younger and older adults [35]. Although differences in dietary intake may contribute to variability in hydrogen levels, the absence of group differences before sleep and the emergence of differences only after sleep suggest that dietary factors alone are unlikely to fully explain the observed age-related differences in ΔH2.
Another possible explanation involves hydrogen consumption related to oxidative processes during sleep. Molecular hydrogen has been reported to act as a selective antioxidant [2], and oxidative stress is known to increase with aging [16]. In addition, sleep and sleep disruption have been associated with increased oxidative stress and metabolic demand [36, 37]. Therefore, it is plausible that hydrogen consumption may be enhanced during sleep in older adults, particularly in the context of fragmented sleep and increased nocturnal awakenings observed in this study.
However, this interpretation remains speculative, as breath hydrogen is an indirect measure and does not directly reflect systemic hydrogen utilization. Further studies incorporating direct markers of oxidative stress will be required to clarify this mechanism.
This study has several limitations. First, dietary intake prior to sampling was not standardized, which may have influenced hydrogen production. Second, bowel habits were not directly assessed, and gastrointestinal status was evaluated only using the CMI. The CMI captures general gastrointestinal complaints but does not provide detailed information on stool frequency or form. Third, breath samples were collected at home and analyzed after transport, with a storage period of approximately 10–12 h, which may introduce variability related to gas diffusion and sample stability [27–29]. Although previous studies suggest that hydrogen remains relatively stable under similar conditions, some degree of measurement variability cannot be excluded. In addition, dietary intake, gastrointestinal motility, concomitant medications, and other factors that may influence gastrointestinal function were not comprehensively controlled. These factors should be addressed in future studies.
In summary, older adults exhibited significantly lower breath hydrogen levels after waking despite similar levels before sleep, indicating age-related differences in overnight hydrogen dynamics. Taken together, these findings suggest that the observed difference in ΔH2 is unlikely to be explained by a single factor such as diet or methane production, but rather reflects a multifactorial process involving gut microbial activity, sleep-related physiology, and hydrogen metabolism. Breath hydrogen measurement may therefore serve as a simple, noninvasive marker for assessing physiological processes associated with sleep and aging.
We thank all the volunteers who participated in this study. We also would like to express special thanks to Ms. Noriko Inui, Mariko Mori, Azusa Kawamura, Shoko Nagahiro, Mariko Komatsu, Nobue Nishi, Hiroko Hayashi, and Yoshiko Kokusho for their technical assistance.
We would like to thank Editage (www.editage.com) for English language editing.
The data cannot be made publicly available upon publication because they contain sensitive personal information. The data that support the findings of this study are available upon reasonable request from the authors.
This study was supported in part by Grants-in-Aid from the Japanese Ministry of Education, Science, and Culture (Grants 24K19512 to Okumura 15J08579 and 19K21435 to Taniguchi).
This study was approved by the Ethics Committee at the National Cerebral and Cardiovascular Research Center (M18-19-2, M26-158) and Chubu University (280031). All participants provided written informed consent before participation in this study. The study was conducted in accordance with the principles of the Declaration of Helsinki.
Comments (0)