
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Remember me
High quality exhaled volatile breath studies require intersubjective reproducible measurements for the results to be trusted. This can only be achieved when:
(i)
the various body regions are in a state of equilibrium according to Henry’s law, a law that demonstrates that a volatile’s concentration in a liquid (e.g. blood) or in tissue (e.g. fat or muscle) is directly proportional to its partial pressure in the headspace above that liquid or in the liquid above the tissue (see appendix A);
(ii)
a steady state is reached in the lungs, i.e. end-tidal exhaled volatile breath concentrations, blood flow, and respiratory flow are all constant (averaged over at least one breath cycle);
(iii)
any changes in cardiac output, alveolar ventilation, and mixed-venous volatile blood concentrations during the collection of exhaled breath samples are taken into consideration;
(iv)
the effects of any inhaled volatiles are correctly accounted for; and
(v)
contributions of volatiles not derived from the alveoli in the lungs are minimal (e.g. minimizing volatiles coming from bacteria in the oral cavity).
Conditions for (i) and (ii) can be achieved by adopting protocols whereby the volunteers providing exhaled breath samples are rested for a given period of time before breath samples are collected. There are currently no standards for this, but approximately 15 min in a seated position should be sufficient to avoid movement related changes in the volatilome. Volunteers should remain calm and still throughout the entire exhaled breath sampling procedure and should breathe in a regular pattern during the collection period. Ideally, the inhaled air should be free from volatiles of interest. When that is not the case, the simple method adopted of subtracting the background inhaled air volatile concentrations from those measured in exhaled breath often leads to incorrect determination of the real endogenous concentrations. The appropriate treatment of this problem is complex, as will be elaborated later. Finally, studies that collect and analyze oral breath samples (either mixed-expired or end-tidal) for volatile analysis, and particularly for use in volatile fingerprinting, e.g. to aid in the diagnoses of diseases, may lead to an incorrect interpretation of the measurements if the microbial volatiles are not identified.
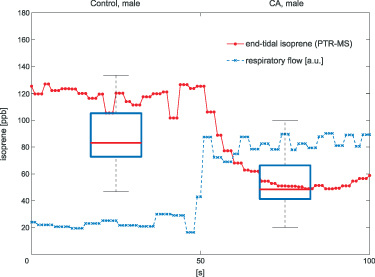
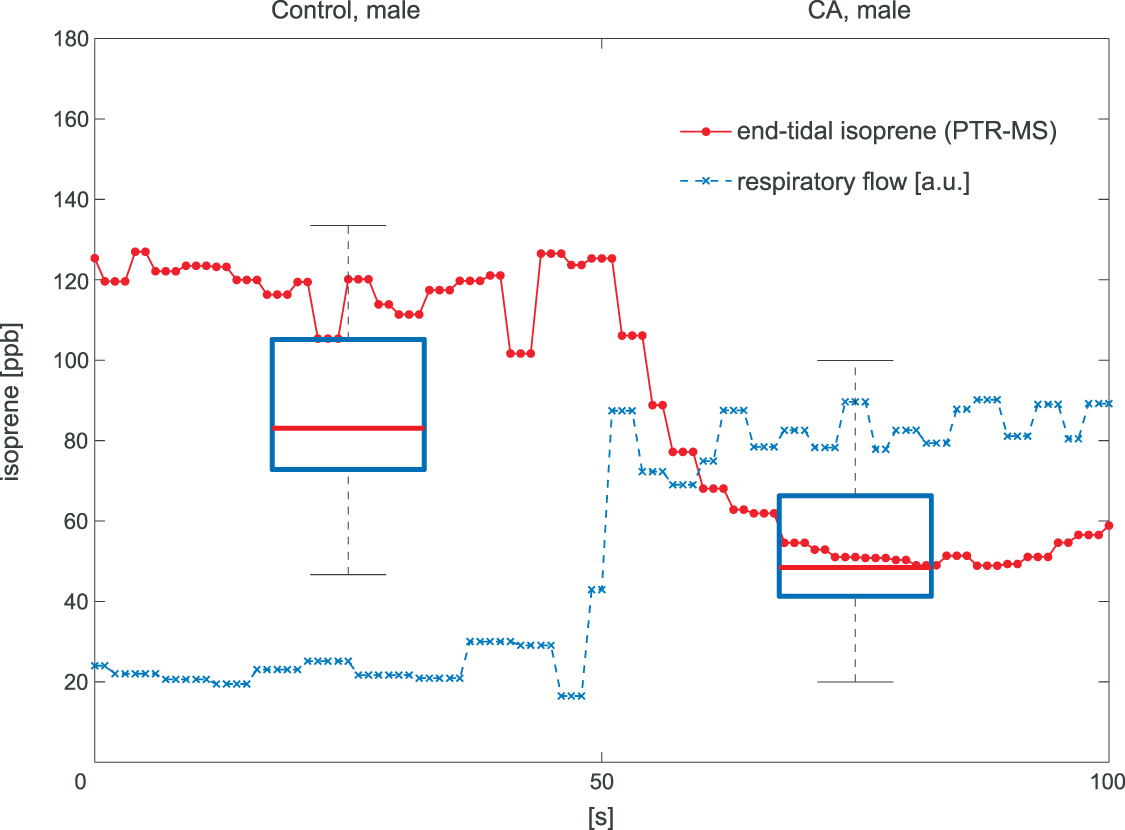
To provide a simple illustrative case of how false interpretations of volatile breath data can occur if a confounding factor is ignored, we demonstrate here the effects of changes in alveolar ventilation during breath sampling. Figure 1 (taken from Koc et al [1]) shows the overlay of results for exhaled isoprene concentrations obtained from two separate and independent studies, with one study investigating lung cancer [2], and the other study examining the effects of hyperventilation [1].
Figure 1. The two box plots show the results obtained for the exhaled breath concentrations of isoprene from breath samples collected from healthy male controls (Control, male) and from late-stage lung cancer male patients (CA, male) (Bajtarevic et al [2]), demonstrating on average much lower exhaled isoprene levels in the breath of the lung cancer patients compared to the healthy controls. These two box plots are overlayed with the results from real-time measurements of exhaled isoprene concentrations (red line with data points) recorded for a healthy male volunteer breathing normally for the first 50 s, followed by hyperventilation for next 50 s, which is illustrated in the figure through the increase in respiratory flow (blue dashed line given in arbitrary units). It can be seen that within about ten seconds of hyperventilating, the healthy volunteer has isoprene levels comparable to those found for lung cancer patients.
Download figure:
Standard image High-resolution imageThe lung cancer study by Bajtarevic et al [2] consisted of analyzing the exhaled isoprene concentrations contained in mixed-expired breath samples collected from 140 males suffering from lung cancer and from 170 healthy (i.e. no lung disease) control males. A summary of the results for that study is provided by the two box plots in figure 1, for which there is a clear distinction between the two volunteer groups (p < 0.01). Given the large number of breath samples taken and analyzed, the results show that there is a statistically significant difference in isoprene concentrations, with the lung cancer patients clearly having on average lower isoprene concentrations than for the healthy volunteers. In agreement with Bajtarevic et al, other studies have also found a reduction of the isoprene concentration in the exhaled breath of patients suffering from lung cancer. For example, recently in a 2024 study by Cheng et al [3] it was found that lung cancer patients had expired isoprene levels below 40 ppb, whereas healthy volunteer isoprene levels were above 60 ppb. (Note that the exhaled concentration values recorded by Cheng et al do not agree with those of Bajtarevic et al, with the levels for the healthy volunteers being in the lung cancer region of Bajtarevic et al study, which must result from issues with differences in calibration). From such studies, it is compelling to conclude that isoprene can be used as a biomarker for lung cancer. However, even if a lower concentration of isoprene in the exhaled breath of lung cancer patients is correct, considerable doubt on the usefulness of isoprene as a biomarker for lung cancer comes from the results of continuous exhaled isoprene measurements in real-time taken over a period of 100 s by Koc et al [1]. Koc et al demonstrate that the exhaled isoprene concentrations from a healthy volunteer can be rapidly reduced by simply changing the breathing pattern. Figure 1 shows that when a healthy volunteer deliberately started to hyperventilate 50 s after normal breathing (blue dashed line), a reduction in exhaled isoprene levels occurred. Given that people with any lung disease are likely to have different breathing patterns compared to healthy people, the observed decrease in isoprene levels for lung cancer patients may not be directly associated with the disease itself, but could simply be a consequence of the effects of the disease on breathing patterns. This illustrative example serves to show that considerable care must be taken in assigning any endogenous volatile to a given disease state when the breathing pattern (e.g. changes in ventilation rates) is not considered, especially for those volatiles with low blood:air partition coefficients (defined by 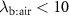 , see [4]), such as isoprene. A mechanistic explanation for this is given by the Farhi equation.
, see [4]), such as isoprene. A mechanistic explanation for this is given by the Farhi equation.
In a 1986 review, Wilson examined the physiological basis and sampling techniques for breath analysis and drew attention to various factors that may affect a volatile’s blood to breath concentration ratio [5]. Wilson emphasized the development of mathematical models to adequately understand the variations of such ratios that are experimentally observed for exhaled breath volatiles and highlighted the importance of the application of the Farhi equation within that modeling. It is therefore appropriate to first discuss the Farhi equation and the consequences of it for exhaled breath volatile analysis. The importance of the Farhi equation is that it relates the alveolar volatile concentration 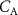 to the mixed-venous blood volatile concentration
to the mixed-venous blood volatile concentration 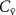 , which is ultimately what we are after because
, which is ultimately what we are after because 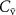 provides a direct window into endogenous metabolism and disease processes within the human body.
provides a direct window into endogenous metabolism and disease processes within the human body.
The derivation of the Farhi equation (see appendix B and [6]) follows from a mass balance differential equation for the lungs, modeled as a single pulmonary compartment and assumes:
(i)
a stationary state is achieved within the lungs, i.e.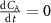 ;
;(ii)
a concentration equilibrium in the alveoli according to Henry’s law, i.e. the volatile organic compound (VOC) of interest forms a simple solution in blood so that Henry’s law is valid, which means that there is no chemical binding of any VOC to blood proteins; and
(iii)
that there is no inhaled concentration, i.e.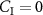 .
.The resulting Farhi equation is:
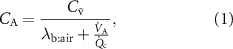
where 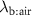 is the blood:air partition coefficient (a dimensionless Henry constant) and
is the blood:air partition coefficient (a dimensionless Henry constant) and 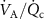 is the alveolar ventilation-perfusion ratio (respiratory flow rate over cardiac output), which is approximately one at rest. In general, the blood:air partition coefficient is a complex parameter resulting from two processes that occur in the blood: partitioning (solubility) and binding [7]. The former is associated with the composition of plasma such as water, lipids and phospholipids content and erythrocytes. The binding, in turn, is determined by plasma proteins’ types and hemoglobin. The partitioning fraction of
is the alveolar ventilation-perfusion ratio (respiratory flow rate over cardiac output), which is approximately one at rest. In general, the blood:air partition coefficient is a complex parameter resulting from two processes that occur in the blood: partitioning (solubility) and binding [7]. The former is associated with the composition of plasma such as water, lipids and phospholipids content and erythrocytes. The binding, in turn, is determined by plasma proteins’ types and hemoglobin. The partitioning fraction of 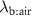 is not expected to change with concentration. However, the binding process exhibits saturation and does change with the concentration of a given volatile. For example, depending on its concentration in blood, approximately 35% to 80% of benzene is bound to plasma proteins [8].
is not expected to change with concentration. However, the binding process exhibits saturation and does change with the concentration of a given volatile. For example, depending on its concentration in blood, approximately 35% to 80% of benzene is bound to plasma proteins [8].
Despite the simplicity of the single compartmental model, the Farhi equation yields valuable insights into the exhalation kinetics of a VOC depending on how 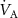 ,
, 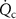 , and
, and 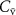 change, as well as showing the dependence of
change, as well as showing the dependence of 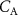 on the blood:air partition coefficient for that VOC, for which there may be individualistic differences, i.e. biological variations and changes associated with diet, medication, age, health, etc but these have never been explored and hence cannot be discussed in this review.
on the blood:air partition coefficient for that VOC, for which there may be individualistic differences, i.e. biological variations and changes associated with diet, medication, age, health, etc but these have never been explored and hence cannot be discussed in this review.
A critical methodological question is the extent to which the measured end-tidal concentration of an exhaled VOC reflects its true alveolar concentration. Can we always assume that 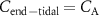 ? Although the simple answer to this question is no; the reasons behind this need an understanding of the solubilities of each and every individual VOC of interest. This is because the requirement for
? Although the simple answer to this question is no; the reasons behind this need an understanding of the solubilities of each and every individual VOC of interest. This is because the requirement for 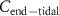 to be equal to
to be equal to 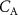 is that the upper airways do not influence the exhaled VOC coming-up from the lungs. This is only true for those VOCs that have low solubilities, a measure for which is given by their blood:air partition coefficients. Generally, it is safe to assume that for volatiles with
is that the upper airways do not influence the exhaled VOC coming-up from the lungs. This is only true for those VOCs that have low solubilities, a measure for which is given by their blood:air partition coefficients. Generally, it is safe to assume that for volatiles with 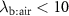 (defined here as low solubility), the effects of the upper airways can be ignored and then it is correct to take
(defined here as low solubility), the effects of the upper airways can be ignored and then it is correct to take 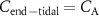 . Examples of VOCs that have
. Examples of VOCs that have 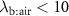 are n-butane [9], ethane [10], isoprene [11], n-pentane [12], and sevoflurane [13], and therefore their end-tidal and alveolar concentrations are the same, provided that there is no inhalation of those volatiles (see section 3).
are n-butane [9], ethane [10], isoprene [11], n-pentane [12], and sevoflurane [13], and therefore their end-tidal and alveolar concentrations are the same, provided that there is no inhalation of those volatiles (see section 3).
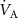 and cardiac output
and cardiac output 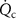 on the alveolar VOC concentrations according to the Farhi equation
on the alveolar VOC concentrations according to the Farhi equationThe Farhi equation (equation (1)) shows that changes in the alveolar concentration 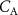 of any given VOC will occur if the ratio
of any given VOC will occur if the ratio 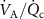 changes, but by how much depends on the blood:air partition coefficient of a VOC. At rest and in a seated position this ratio is approximately one. Whilst during exercise, hyperventilation (predominantly increasing
changes, but by how much depends on the blood:air partition coefficient of a VOC. At rest and in a seated position this ratio is approximately one. Whilst during exercise, hyperventilation (predominantly increasing 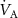 ), or during times of stress, this ratio increases, it still remains a low value (generally less than 4) and hence any change will only have an influence on the alveolar concentrations
), or during times of stress, this ratio increases, it still remains a low value (generally less than 4) and hence any change will only have an influence on the alveolar concentrations 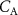 for those VOCs with blood:air partition coefficients less than about 10. Such effects have been observed for, e.g. isoprene, methane, and n-butane [14, 15].
for those VOCs with blood:air partition coefficients less than about 10. Such effects have been observed for, e.g. isoprene, methane, and n-butane [14, 15].
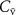 on the alveolar concentration according to the Farhi equation
on the alveolar concentration according to the Farhi equationChanges in the mixed venous volatile concentration 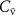 occur when the relative blood flows to different body regions change (e.g. as a result of physical exercise) and/or when variations occur in the endogenous production of that volatile. Physical exercise has been thoroughly examined for isoprene. The sudden increase in blood flow through muscle tissue (where the production of isoprene in the body occurs [16, 17]) during exercise results in a higher concentration of isoprene in the mixed venous blood for a given period of time [18, 19]. This period of time relates to the washout of isoprene from the muscle tissue, the production of which cannot keep-up with the loss so that a new steady-state is reached. This explains the observed rapid increase and then the following decrease in isoprene levels in mixed-venous blood just after the start of the exercise and during the exercise, respectively. This is in agreement with expectations from the Farhi equation due to the increase in
occur when the relative blood flows to different body regions change (e.g. as a result of physical exercise) and/or when variations occur in the endogenous production of that volatile. Physical exercise has been thoroughly examined for isoprene. The sudden increase in blood flow through muscle tissue (where the production of isoprene in the body occurs [16, 17]) during exercise results in a higher concentration of isoprene in the mixed venous blood for a given period of time [18, 19]. This period of time relates to the washout of isoprene from the muscle tissue, the production of which cannot keep-up with the loss so that a new steady-state is reached. This explains the observed rapid increase and then the following decrease in isoprene levels in mixed-venous blood just after the start of the exercise and during the exercise, respectively. This is in agreement with expectations from the Farhi equation due to the increase in 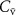 . For methane, an opposite effect is observed, whereby the mixed venous blood concentration decreases during exercise. This is because methane in the human body comes from anaerobic fermentation by methanogens predominantly in the large intestine. This methane that originates in the gut traverses the intestinal mucosa to be absorbed into the systemic circulation. During exercise, the relative blood flow to the intestine is reduced. Together with an increase in the ratio
. For methane, an opposite effect is observed, whereby the mixed venous blood concentration decreases during exercise. This is because methane in the human body comes from anaerobic fermentation by methanogens predominantly in the large intestine. This methane that originates in the gut traverses the intestinal mucosa to be absorbed into the systemic circulation. During exercise, the relative blood flow to the intestine is reduced. Together with an increase in the ratio 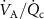 , by about a factor of 2 during exercise, and the decrease in fractional blood flow to the gut, by also nearly a factor of 2, this leads to the observed decrease in exhaled methane by approximately a factor of 4 during exercise, again as expected from the Farhi equation [20].
, by about a factor of 2 during exercise, and the decrease in fractional blood flow to the gut, by also nearly a factor of 2, this leads to the observed decrease in exhaled methane by approximately a factor of 4 during exercise, again as expected from the Farhi equation [20].
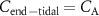 ?
?For end-tidal concentrations of an exhaled VOC to be the same as the alveolar values, i.e. 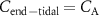 , certain conditions are needed:
, certain conditions are needed:
Under these conditions, the Farhi equation can be written as:
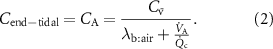 2.4. What happens when
2.4. What happens when 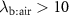 ? Is it safe to use the Farhi equation?
? Is it safe to use the Farhi equation?The simple answer to these two questions is that the Farhi equation is valid for all VOCs, but in the case of extremely high blood:air partition coefficients, e.g. acetone at 340, changes in ventilation and perfusion will have a negligible effect on 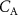 given that the range of
given that the range of 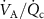 is anywhere between approximately 1 and 4, depending on whether a person is at rest or not. Thus, for a VOC with a high blood:air partition coefficient, the Farhi equation reduces to:
is anywhere between approximately 1 and 4, depending on whether a person is at rest or not. Thus, for a VOC with a high blood:air partition coefficient, the Farhi equation reduces to:
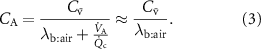
Therefore, changes in the 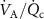 should not have an effect on the observed exhaled levels, and therefore end-tidal exhaled breath concentrations of such VOCs are expected to show no change if we assume that they are equal to the alveolar concentration. However, this is not valid for volatiles with sufficiently high blood:air partition coefficients for which the end-tidal exhaled concentration is equal to that in the bronchi rather than that in the alveoli:
should not have an effect on the observed exhaled levels, and therefore end-tidal exhaled breath concentrations of such VOCs are expected to show no change if we assume that they are equal to the alveolar concentration. However, this is not valid for volatiles with sufficiently high blood:air partition coefficients for which the end-tidal exhaled concentration is equal to that in the bronchi rather than that in the alveoli:
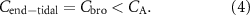
The difference between 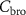 and
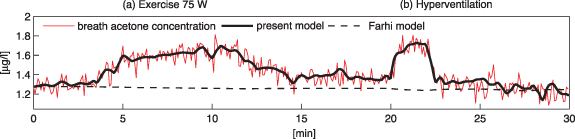
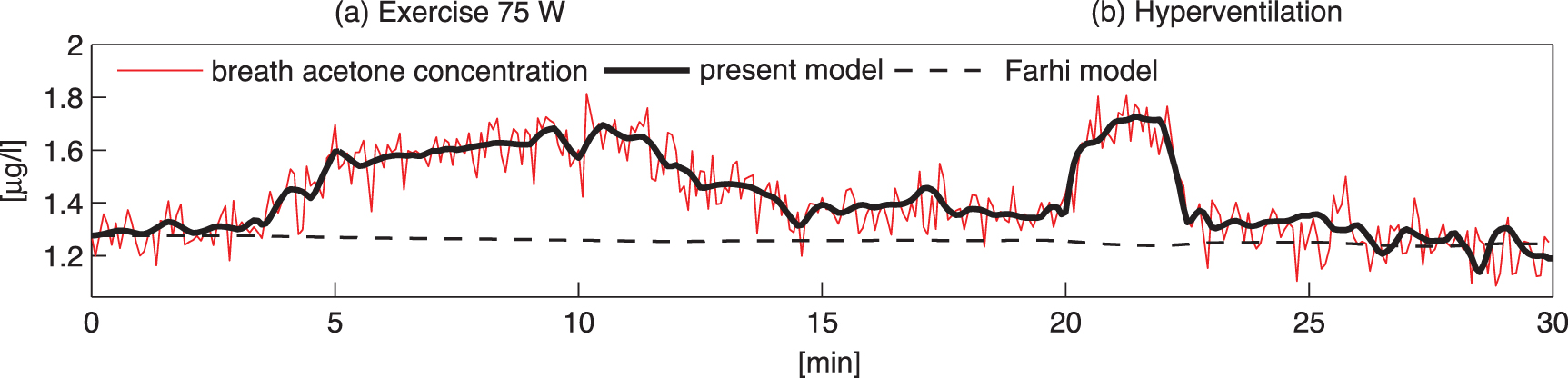
and 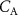 is due to absorption and desorption processes in water-like mucosa lining the upper airways, which are more dominant for VOCs with higher solubilities. Multiple studies with ethanol and acetone have confirmed such effects [4, 21, 22]. For perspective, figure 2 shows end-tidal exhaled acetone levels in real-time first during an exercise period, then secondly a period of hyperventilation with increased tidal volume, and then finally hyperventilation at high breathing frequency. For the exercise period and the first hyperventilation period, sudden increases in acetone concentrations are observed [21]. These increases in acetone concentrations result from increases in air flow rate from the lungs, leading to more acetone being drawn up from the alveoli into the upper airways, which in turn results in a new higher end-tidal exhaled acetone concentration. For the second hyperventilation phase, due to the higher frequency of breathing, the air flow remains mainly in the upper airways, so that no noticeable additional acetone is drawn-up from the alveoli.
is due to absorption and desorption processes in water-like mucosa lining the upper airways, which are more dominant for VOCs with higher solubilities. Multiple studies with ethanol and acetone have confirmed such effects [4, 21, 22]. For perspective, figure 2 shows end-tidal exhaled acetone levels in real-time first during an exercise period, then secondly a period of hyperventilation with increased tidal volume, and then finally hyperventilation at high breathing frequency. For the exercise period and the first hyperventilation period, sudden increases in acetone concentrations are observed [21]. These increases in acetone concentrations result from increases in air flow rate from the lungs, leading to more acetone being drawn up from the alveoli into the upper airways, which in turn results in a new higher end-tidal exhaled acetone concentration. For the second hyperventilation phase, due to the higher frequency of breathing, the air flow remains mainly in the upper airways, so that no noticeable additional acetone is drawn-up from the alveoli.
Figure 2. A real-time profile of measured end-tidal (bronchial) exhaled acetone concentrations (red line) exhaled by a volunteer on an exercise bicycle. The observed changes in the measured acetone concentrations are in response to the following physiological regime: volunteer at complete rest (0–3 min), exercise on a bicycle at 75 W (3–12 min), another complete rest (12–20 min), hyperventilation with increased tidal volume (20–22 min) whilst doing no exercise, rest (22–25 min), high-frequency hyperventilation (25–28 min) whilst doing no exercise, and finally another period of complete rest (28–30 min). Superimposed on the acetone measurements are the results obtained from the modeling for the bronchial concentrations (continuous black line). This figure is adapted from figure 6 of [21], with permission from Springer Nature, to which the reader is referred to for a more in-depth description.
Download figure:
Standard image High-resolution imageIn conclusion, the measurable end-tidal (bronchial) breath concentration of a highly soluble VOC is generally lower than the corresponding alveolar concentration, and by how much depends on a number of physiological parameters that are difficult to quantify experimentally, such as airway temperature (affecting solubility) or fractional bronchial perfusion, see equation (21) in [21]. In order to determine more reliable alveolar and mixed-venous concentrations for volatiles with 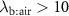 isothermal re-breathing methods (equilibrating
isothermal re-breathing methods (equilibrating 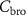 and
and 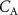 [23, 24]) could be employed. However, as discussed later, isothermal re-breathing protocols are not practical under most breath sampling scenarios. As an alternative to isothermal re-breathing methods, the end-tidal breath concentration of a volatile could be standardized to a reference respiratory gas, such as
[23, 24]) could be employed. However, as discussed later, isothermal re-breathing protocols are not practical under most breath sampling scenarios. As an alternative to isothermal re-breathing methods, the end-tidal breath concentration of a volatile could be standardized to a reference respiratory gas, such as 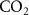 ,
, 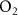 , or water vapor [25].
, or water vapor [25].
Volatiles that are found in exhaled breath have endogenous (systemic) and/or exogenous origins. Endogenous volatiles result from normal and abnormal metabolic bodily processes or from pathological disorders, i.e. they originate within the body. Exogenous volatiles found in exhaled breath result from volatiles introduced into the human body from external sources, e.g. by ingestion, inhalation, or dermal exposure. Whilst volatiles found in exhaled breath can have contributions from both endogenous production and external sources, a subgroup of exogenous volatiles found in breath are completely foreign to the human body, i.e. they do not result from endogenous processes, there are no endogenous contributions to the exhaled breath. These are referred to as xenobiotic volatiles. Importantly, such compounds are often lipophilic, which means that they will accumulate in fat tissue if they are not rapidly metabolized in the human body. This has been shown to occur for ingested limonene, a xenobiotic volatile, that has considerable potential for use as a biomarker for chronic liver disease [26–29]. Thus, xenobiotic volatiles may not only be confounders in exhaled volatile analysis, but they can also act as potential biomarkers for disease due to changes in normal metabolic processes.
Volatiles present in inhaled air constitute a critical confounding factor in interpreting exhaled breath VOCs. The effects of this can be minimized by adopting appropriate mathematical procedures and breath sampling methods. For breath collection, exhaled samples should not be taken in rooms/locations containing VOC sources, such as chemicals, disinfectants, cleaning agents, drugs, etc that are commonly present, e.g. in a clinical environment. However, this is not always practical or possible. In such cases, when a VOC under study is present in the room air, to accurately interpret its exhaled breath concentration the influence of the inhaled volatile concentration needs to be taken into account. The correct handling of the inhaled room air volatiles to determine the correct exhaled breath volatile levels has been a major issue of debate within the breath research community (see, e.g. [30–32] and the reviews [33, 34]).
In 1999 Phillips et al [30] summarized the situation by stating that ‘researchers have responded to this dilemma with three different strategies’. These three strategies are:
(1)
to ignore the problem;
(2)
to provide the subject with VOC-free air to breathe prior to collection of the breath sample; or
(3)
to correct for the problem by subtracting the background VOC concentrations measured in room air from the measured concentrations of the same volatiles exhaled.
Phillips et al selected the third option, which they called the ‘alveolar gradient’ method. However, a simple subtraction can sometimes lead to negative concentration values, which physically does not make sense, and hence already indicates that this method is too simple. This apparent mathematically negative concentration value will always occur when the inhaled room air concentration of a volatile is greater than the exhaled concentration. The alveolar gradient method only works for those volatiles that once inhaled are not stored, metabolized within, and/or eliminated from the body (other than through exhalation). The inhaled concentration then simply adds to the exhaled concentration, i.e. what is breathed in will be exhaled out. One such example is methane. However, for the majority of endogenous volatiles this is not the case, and hence the alveolar gradient approach requires adjustment to yield the correct endogenous exhaled concentrations, which are directly correlated to the associated mixed-venous blood volatile concentrations.
The conclusive evidence for this adjustment of the alveolar gradient method comes from a research study by Spanel et al [35]. They investigated the dependence of exhaled volatile concentrations on their inhaled volatile concentrations for a number of common endogenously produced volatiles, namely: acetone, ammonia, isoprene, methanol, and pentane. In addition to these endogenous volatiles, Spanel et al investigated formaldehyde. For each of these VOCs, they determined that the exhaled breath volatile concentration shows a linear relationship to the inhaled volatile concentration (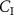 ), i.e.
), i.e. 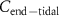 is directly proportional to
is directly proportional to 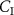 .
.
Modeling has shown that this empirically observed linear relationship is correct, provided that the cardiac output and ventilation during the breath sampling are kept constant [36, 37]. These modeling studies also provide a value for the gradient of the linear slope that is needed to determine the fraction of the inhaled volatile to be subtracted from the exhaled values. This value depends very much on the volatile’s blood:air partition coefficient, the magnitude of which determines how important other physiological factors affect the impact of the inhaled volatile concentrations.
3.1. Room air correction for VOCs with blood:air partition coefficient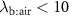
As described in the previous section, 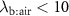 means that the measured end-tidal VOC concentrations are one and the same as the alveolar concentrations
means that the measured end-tidal VOC concentrations are one and the same as the alveolar concentrations 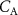 , and at rest the exhalation kinetics are accurately described by the Farhi equation (equation (1)). To this end, a simple two-compartmental model that generalizes the Farhi equation to the case where the inhaled concentration
, and at rest the exhalation kinetics are accurately described by the Farhi equation (equation (1)). To this end, a simple two-compartmental model that generalizes the Farhi equation to the case where the inhaled concentration 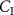 of a VOC is not zero was developed by Unterkofler et al [36]. In addition to a lung compartment, it includes a body compartment to account for volatile metabolism and production. The modeling determines that the relationship of
of a VOC is not zero was developed by Unterkofler et al [36]. In addition to a lung compartment, it includes a body compartment to account for volatile metabolism and production. The modeling determines that the relationship of 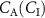 to
to 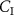 is:
is:
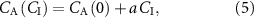
where the gradient a is given by
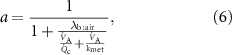
and the intercept that provides the correct alveolar concentration, i.e. with no contribution from an inhaled volatile, is given by:
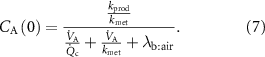
See appendix C and [36] for derivations of the two above equations. As can be seen, the end-tidal concentration 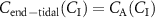 (
(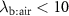 ) depends not only on the inhaled concentration, but also on various physiological parameters, namely the blood:air partition coefficient, the ventilation and perfusion rates (
) depends not only on the inhaled concentration, but also on various physiological parameters, namely the blood:air partition coefficient, the ventilation and perfusion rates (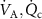 ), the volatile production rate (
), the volatile production rate (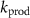 ), and the volatile metabolic rate (
), and the volatile metabolic rate (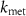 ). This reveals that in order to obtain the true alveolar volatile concentration
). This reveals that in order to obtain the true alveolar volatile concentration 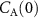 ,
, 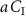 must be subtracted from the measured value, i.e.
must be subtracted from the measured value, i.e.
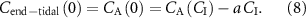
For comparison, note that the alveolar gradient method claims that a = 1 for all VOCs, thus neglecting the physiological and physico-chemical influences in equation (6). The proportion 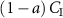 represents that fraction of the inhaled concentration that is eliminated (not through exhaled breath), stored, and/or metabolized within the body.
represents that fraction of the inhaled concentration that is eliminated (not through exhaled breath), stored, and/or metabolized within the body.
This model was tested and validated using real-time exhalation data for a volunteer at rest or on an exercise bicycle whilst maintaining 75 W for given periods of time and whilst inhaling deuterated isoprene-d5 [36]. This volunteer was breathing freely through a mask to which an analytical soft chemical ionization mass spectrometric instrument was directly connected to record the inhaled and exhaled isoprene-d5 concentrations in real-time. Figure 3(a) shows the recorded endogenous isoprene behaves as expected, i.e. isoprene levels of about 100 ppb whilst the volunteer was at rest and then sudden increases in concentrations are observed at the start of the exercise. Since the exhaled isoprene originates from production in muscle tissue, the increased blood flow through the muscle tissue during the exercise phase leads to increases in the mixed venous isoprene blood concentrations, which in turn leads to the observed peak in the exhaled breath isoprene concentration soon after the start of the exercise. This is followed by a decrease in the exhaled isoprene concentration until a new stationary state (between increased blood flow and production) is reached in the muscle tissue. After a period of rest, the beginning of the second exercise session shows a lower second isoprene peak. This is because the muscle tissue has not produced sufficient isoprene in the rest period to compensate for what has been lost from the original steady state [16]. In contrast, following the release of isoprene-d5 into the exercise room in minute nine, the inhaled isoprene-d5 is distributed rapidly and evenly throughout the body and no concentration peak is observed at the beginning of the exercise. Instead, a drop in exhaled isoprene-d5 concentration can be seen, as expected from the Farhi equation due to a delay in the transport of isoprene-d5 throughout the body. However, this is then compensated by increased inhalation during exercise, which ultimately leads to a higher exhaled concentration (red line) compared to the exhaled concentration at rest (black line). Quantitatively, this can be explained by a higher value of the gradient, a, when transitioning from rest to exercise, as expected from equation (6) due to alveolar ventilation 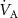 increasing by approximately a factor of four. At the end of the exercise study, when the mask was removed from the volunteer, the volatile concentrations in the room were measured through the mask that is directly connected to the analytical instrument. The slow decrease in the concentration of isoprene-d5 in the room during the course of the measurements resulted from losses of the compound because the laboratory was not airtight.
increasing by approximately a factor of four. At the end of the exercise study, when the mask was removed from the volunteer, the volatile concentrations in the room were measured through the mask that is directly connected to the analytical instrument. The slow decrease in the concentration of isoprene-d5 in the room during the course of the measurements resulted from losses of the compound because the laboratory was not airtight.
Figure 3. Real-time end-tidal isoprene concentration measurements for a volunteer at rest and during two exercise periods (75 W) for endogenous isoprene (top panel) and (b) inhaled isoprene-d5 (bottom panel). Figure adapted from [36]. © IOP Publishing Ltd. CC BY 3.0.
Download figure:
Comments (0)