
Remember me
The inflammatory bowel disease (IBD), including Crohn’s disease (CD) and ulcerative colitis (UC), is a group of inflammatory disorders of the gastrointestinal tract with an unknown etiology. The prevalence of IBD is approximately 0.3% of the European population and has significantly increased in recent years. IBD primarily manifests as diarrhea and abdominal pain; however, over the course of the disease, several serious complications may develop, including colorectal cancer (CRC). IBD-associated CRC, or colitis-associated CRC (CAC), is a form of CRC arising in IBD patients. CAC accounts for approximately 1–2% of all CRC cases and is characterized by distinct clinical features and pathogenesis. The overall incidence of CRC in IBD patients is estimated at 1%, 1.5%, and 2.7% at 10, 20, and 30 years after diagnosis; however, these data vary among studies [1]. Notably, a relatively small fraction of IBD patients develop CAC, but it constitutes approximately 10–15% of deaths among these patients [2]. The exact pathogenesis of CAC remains obscure. Consequently, molecular biomarkers such as albumin levels, DNA, RNA, proteins, and miRNAs have recently been investigated to improve diagnostic modalities, prognostic assessment, and therapy [3,4,5].
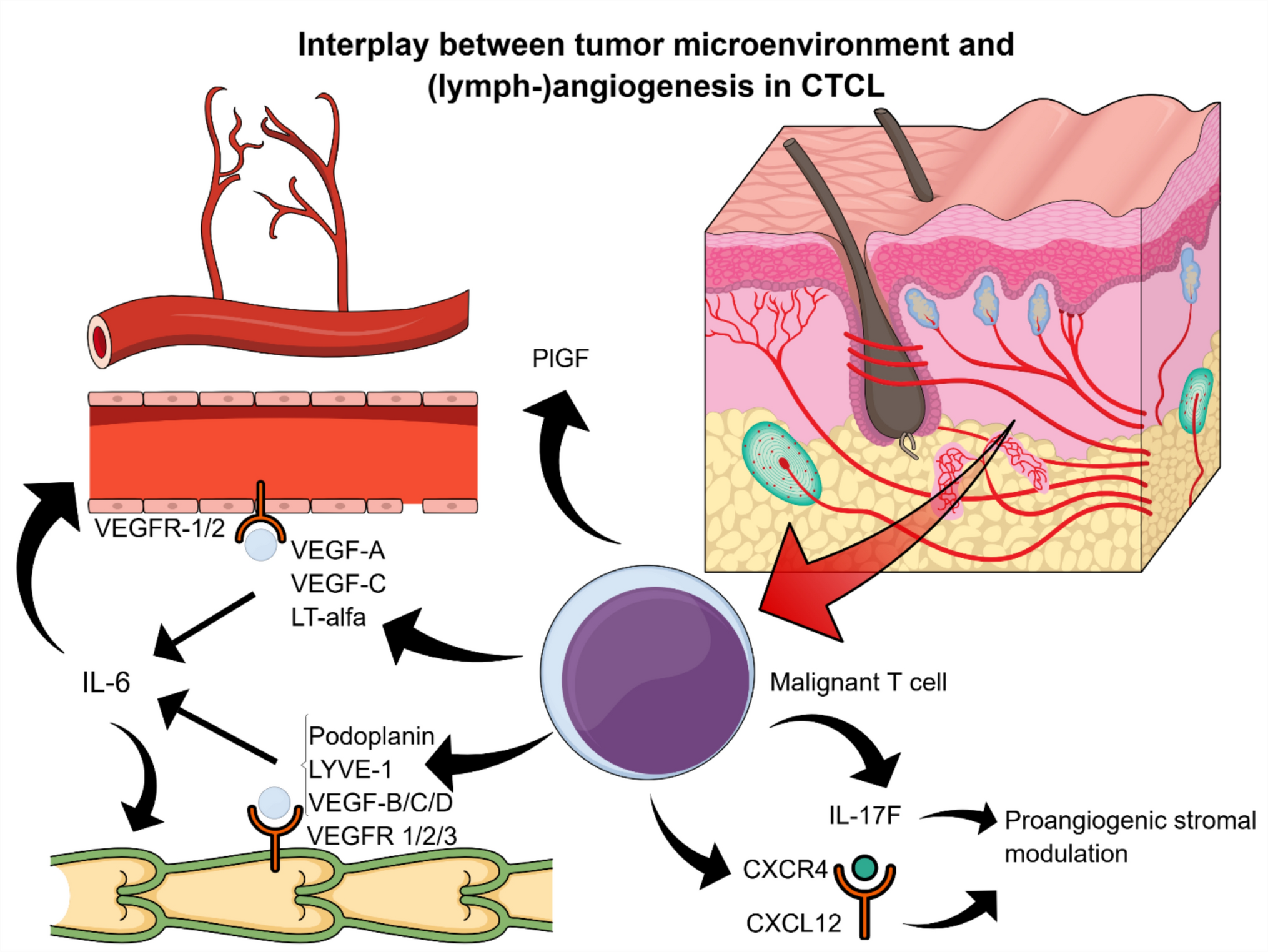
MicroRNAs (miRNAs) are a group of noncoding, single-stranded RNA biomolecules that usually bind to the 3’-untranslated region (3ʹ-UTR) of their specific target mRNAs, thereby suppressing protein synthesis posttranscriptionally (Fig. 1). First discovered in the 1990s and recognized as a class of biological regulators in the 2000s, miRNAs have recently gained popularity as tools for diagnosis and therapy [6]. Multiple studies suggest promising results involving miRNAs in CRC and IBD patients, which could lead to significant changes in clinical practice [5, 6]. miRNAs control the expression of over a thousand transcripts, either directly by binding or indirectly by targeting transcription factors, epigenetic regulators, or effectors of signal transduction pathways. miRNAs are more stable than mRNAs; therefore, they are easily detectable in formalin-fixed, paraffin-embedded tissues and plasma/serum using reliable methods, including quantitative reverse transcription PCR (qRT-PCR) and microarray analysis. For this reason, miRNAs are highlighted as biomarkers that may be sensitive and specific, thus potentially useful for early detection, prognostic classification of CAC, and therapeutic decision-making in its treatment [6, 7].
Fig. 1
miRNA biogenesis. miRNAs, similar to mRNA, are transcribed by RNA polymerase II (Pol II) as stem-loop primary miRNA (pri-miRNA). The DGCR8 protein recognizes specific sequences, and the DROSHA endonuclease cleaves them, forming the characteristic hairpin structure of precursor miRNA (pre-miRNA). Pre-miRNA is transported through nuclear pores by exportin-5 (XPO5) in association with the RAN protein. In the cytoplasm, a protein complex containing the DICER nuclease and TARBP protein processes the hairpin structure, generating a miRNA duplex composed of a 5ʹ-miRNA strand and a 3ʹ-miRNA strand. Following unwinding of the duplex, one strand—typically the 3ʹ strand—is degraded, while the remaining strand forms the mature miRNA. This mature miRNA associates with proteins, including Argonaute (AGO), to form the RNA-induced silencing complex (RISC). miRNAs associated with RISC recognize short complementary sequences (known as the seed region) in the 3ʹ-UTR of target mRNAs. A single miRNA can bind to multiple transcripts, leading to repression of the translation of the target gene
In this review, we discuss available studies and outline future directions concerning the validity of using miRNAs in diagnosing and treating CAC and CRC, which may interest clinicians, diagnosticians, and scientists involved in developing new diagnostic and therapeutic strategies. This review comprehensively answers questions about which miRNAs, according to the current state of knowledge, may serve as specific biomarkers of CAC and CRC. It also examines their connections with inflammation and epithelial-mesenchymal transition (EMT), and outlines the possibilities of clinical application of this knowledge in the context of diagnosis and treatment of these diseases.
The overview of the key miRNA in CRC and CACIt has been confirmed that infiltrating inflammatory cells and proinflammatory mediators in the intestinal microenvironment play a crucial role in colon carcinogenesis among IBD patients [8]. These inflammatory mediators include cytokines, chemokines, transcription factors, reactive oxygen and nitrogen species, prostaglandins, and miRNAs. The expression of miRNAs is altered in various types of human cancer studied to date, including CAC. Specific miRNAs have been shown to have oncogenic or tumor-suppressive properties, highlighting their role as key factors in carcinogenesis [9].
The scientific literature contains a growing number of reports about miRNAs associated with IBD, CD, CAC, and CRC, some of which are listed in Table 1.
Table 1 miRNA level changes in UC, CD, CRC, and CACHowever, there are still several miRNAs that have not been evaluated in CAC. Nonetheless, their role has been confirmed in CRC and warrants further investigation among IBD patients. miR-217, miR-144, miR-129, miR-125a, miR-125b, miR-375, miR-328, and miR-194 have been identified as useful prognostic biomarkers of CRC [24, 27, 28, 39,40,41]. Other miRNAs that are elevated in CRC include miR-21 and miR-31, both of which are linked to advanced cancer stages [42]. Additionally, miR-155 has been shown to increase in CRC and is thought to play a role in tumor development, intestinal barrier dysfunction, and inflammation-related pathways [43]. Moreover, the downregulation of miR-145, miR-192, and miR-375 has been correlated with CRC [44,45,46]. At the same time, an increase in miR-17-5p leads to a decrease in phosphatase and tensin homolog (PTEN), which is associated with resistance to conventional therapy for CRC [47]. Several miRNAs (miR-21, miR-29a, miR-92a, miR-221, and others) are also believed to be related to different stages of CRC, thus opening up the possibility of a new staging technique in patients [48]. Nevertheless, studies suggest that the pathogenesis of CAC differs from that of CRC; therefore, the exact role of CRC-related miRNAs requires further evaluation. The results of our literature search for miRNAs associated with CAC and CRC, including animal and human studies, are described below and summarized in Table 2.
Table 2 Selected miRNA roles and possible usemiR-21miR-21 is one of the first miRNAs for which an increased expression has been associated with cancer development. Extensive research shows that miR-21 is associated with lung cancer, pancreatic cancer, gliomas, breast and prostate cancer, among others [56,57,58]. miR-21 is also one of the most important miRNAs involved in the progression of CRC [56].
miR-21 levels are higher in CRC and inflammatory disorders, whether the inflammation is due to IBD or acute intestinal obstruction. Evidence suggests that miR-21 impairs intestinal barrier function and increases the secretion of antimicrobial peptides, causing gut dysbiosis and thus acting as a pro-inflammatory agent in IBD [59, 60].
miR-21 is believed to be an archetypal oncogenic factor, found to be upregulated in inflammation, trauma, and CRC patients at advanced stages. By inhibiting nuclear receptor NR2F2 (nuclear receptor subfamily 2 group F member 2) and SMAD7 (Mothers against decapentaplegic homolog 7) expression, miR-21 also promotes TGF-β-induced EMT in CRC, which initiates cancer metastasis and progression [61]. Using miR-21 as a biomarker for CRC has been found effective since its sensitivity was calculated to be 0.77 and specificity to be 0.84; it is also easily detectable [62].
As it is not only present during carcinogenesis but is also actively oncogenetic, miR-21 could potentially be used in therapy. Multiple targets have been identified for miR-21 which show the potential of being modulated to regulate cell proliferation and cancer invasion, such as NR2F2, tumor suppressor PTEN, programmed cell death 4 (PDCD4), Salvador 1 protein (SAV1), RAS homolog family member B (RhoB), cell division cycle 42 (CDC42), and 15-hydroxyprostaglandin dehydrogenase (15-PGDH) [63,64,65]. In particular, the connection between miR-21, PDCD4, and PTEN appears especially promising [66]. Ke et al. [67] also recently demonstrated that miR-21 promotes UC development via TNF-α (tumor necrosis factor α) and changes in the gut microbiota.
Current data regarding the role of miR-21 in CAC pathogenesis primarily stems from animal studies. Shi et al. performed a study on miR-21 knocked-out (KO) mice with AOM/DSS (azoxymethane/ dextran sodium sulfate)-induced CAC, which is 99 accepted experimental model of inflammation-induced CRC. The AOM/DSS mouse model was established by intraperitoneally administering AOS at a dose of 12 mg/kg to 8–10-week-old male mice. Seven days later, the mice were given 2% DSS in their drinking water for five consecutive days. On day 20, a second dose of AOM (12 mg/kg) was given, followed by another 2% DSS treatment for five days, starting seven days after the second AOM injection. Mice were then maintained on plain water and monitored until the end of the experiment, with tumor assessments conducted on days 31 and 81. The study revealed that KO mice, compared with wild-type (WT) mice were characterized by decreased b-catenin levels, increased E-cadherin levels, and reduced expression of Ki67. Furthermore, levels of suppressor protein PDCD4, a potential target of miR-21, were statistically higher in miR-21 KO mice compared with WT mice. Moreover, the authors reported that increased levels of miR-21 were negatively correlated with levels of b-catenin and positively correlated with E-cadherin levels. In summary, miR-21 deficient mice developed less severe neoplastic conditions and appeared resistant to CAC induction [18].
Another study performed by El-Daly et al. [68] revealed that mice with AOM/DSS-induced CAC had higher serum levels of miR-21 compared to the control group. In the same study, higher levels of miR-141, miR-15b, miR-29a, carcinoembryonic antigen, and cancer antigen 19-9 were associated with CAC development. Lai et al. [69] conducted a study on transgenic zebrafish, which revealed that miR-21 is crucial in tumor development in AOM/DSS-induced CAC.
Only a few studies on miR-21 in human CAC are available. Shi et al. demonstrated that UC patients with dysplasia and cancer exhibited significantly higher levels of miR-21 compared to patients with active UC [18]. A similar result was reported by Ludwig K. et al., who analyzed UC patients with and without dysplasia, revealing that miR-21 levels were higher in the former group [15]. Furthermore, Yang et al. [70] revealed that levels of miR-21 increase with the duration of UC. All these studies may help explain the increasing risk of CAC development in the course of IBD.
miR-301amiR-301a plays a role in multiple biological functions that lead to different types of cancer, such as hepatocellular carcinoma, lung cancer, pancreatic ductal adenocarcinoma, breast cancer, and osteosarcoma; it is also present in the development and progression of both, IBD and CRC [49, 50]. miR-301a is proven to suppress apoptosis; it stimulates the proliferation and metastasis of pancreatic, colon, and gastric cancer; it also plays a role in the progression of drug resistance to cancer. Moreover, miR-301a is suspected to control NF-kB, which plays a crucial role in inflammation-driven carcinogenesis [63].
miR-301a is upregulated in inflamed bowel lesions in IBD, as opposed to no sign of the rise of its levels in mucosa tissue in a healthy patient [49]. Moreover, in lung and colon carcinogenesis miR-301a affects tumor-associated immune cells and through that enhances inflammation [71]. There is also a correlation between miR-301a and IL-1β levels, with IL-1β stimulating the expression of miR-301a in CRC cells and overexpression of miR-301a enhancing the expression of IL-1β, along with IL-6, IL-8, and TNF-α. Different studies show that miR-301a is a factor in mucosal permeability development and regulation of IBD and CAC; therefore, it might play a significant role in the disruption of gastrointestinal immunology [72].
The study conducted by He et al. demonstrated that miR-301a-deficient mice were less susceptible to AOM/DSS-induced CRC and developed fewer tumors compared to the control. Furthermore, miR-301a was proposed to upregulate NF-kB levels in a BTG1 (B cell translocation gene 1)-dependent manner. In the same study, samples from IBD patients were collected and analyzed for miR-301a, BTG1, and NF-kB levels. The analysis revealed that both miR-301a and NF-kB levels were elevated in IBD patients, while BTG1 levels decreased. The authors identified BTG1 as a direct target of miR-301a, a finding confirmed by in vitro studies. Since BTG1 is regarded as a negative regulator of NF-κB, its silencing by miR-301a may contribute to enhanced inflammation driven by the NF-κB pathway (nuclear factor kappa-light-chain-enhancer of activated B cells) [36, 73].
Ma et al. performed a similar study, investigating the role of miR-301a and signal transducer and activator of transcription 3 (STAT3) in developing inflammation-induced lung and CRC. In both cases, carcinogenesis was diminished in miR-301a deficient mice. Nearly 50% of subjects did not develop any tumors and the remaining half developed tumors of smaller sizes and better differentiated architecture. This study suggests that control of NF-kB by miR-301 may occur via STAT3 [49]. In addition, Zhang et al. showed that the level of miR-301a-3p is significantly increased in the HT-29 and SW620 cell lines, as well as in the tissues of CRC patients. miR-301a-3p also contributes to tumor progression by negatively regulating deleted in liver cancer-1 (DLC-1) and runt-related transcription factor 3 (RUNX3), which inhibit migration, proliferation, as well as induce apoptosis in vitro models of CRC [74].
Furthermore, a study conducted by Yang et al. [75] revealed that miR-301a was significantly higher in patients with CAC compared with CRC indicating that this miRNA may play a significant role in the pathogenesis of this disease.
miR-31miR-31 is increased in CRC and is associated with cancers characterized by deeper invasion and advanced TNM stages; the level of miR-31 is higher at TNM stages III-IV than at lower stages. miR-31 plays a role in the promotion of cell proliferation, migration, and invasion, as well as the regulation of tumor growth through HIF-1α (hypoxia-inducible factor 1). Overexpression of miR-31 inhibits the tumor suppressor E2F2, leading to increased proliferation of CRC cells [51].
A connection has also been found between the expression of miR-31 and intestinal barrier dysfunction, which directly influences inflammation and cancer development; it regulates colonic inflammation in IBD by targeting HIF-1a, which, apart from inflammation, also controls cell functions and tumor growth [76]. Thus, miR-31 is linked to the promotion of cell proliferation, invasion, and migration. It is also found to directly target the large tumor suppressor kinase 2 (LATS2), thereby modulating the Hippo pathway, which controls organ size. The downregulation of this pathway is associated with carcinogenesis [77].
A major study about the role of miR-31 in IBD and CAC was performed by Olaru et al. A comparison of IBD patients with healthy individuals revealed that miR-31 levels in tissue are statistically higher in the IBD group. Moreover, IBD patients with neoplastic lesions exhibited 11-fold higher levels of miR-31 than nonneoplastic IBD patients. Intriguingly, the authors reported that miR-31 levels were 20-fold higher in CAC patients compared with sporadic CRC [23].
Another study revealed, that upregulation of miR-31 is associated with dysplastic lesions in patients with UC. However, the authors do not describe the possible target of miR-31 [78].
Similar results are reported by Necela et al. The author revealed, that upregulation of miR-31 is associated with tumorigenesis in the course of bowel inflammation and may be useful in distinguishing specimens with chronic colitis from those with neoplastic progression [79].
In turn, Sun et al. identify RAS p21 GTPase activating protein 1 (RASA1) as a direct target of miR-31, involved in the RAS signaling pathway, the regulation of which by this miRNA significantly accelerates the proliferation of CRC cells in in vitro studies [80]. Moreover, recent studies indicate that factors downregulating miR-31 expression may be a good therapeutic target in CRC. Li et al. reported that long noncoding RNA (lncRNA) MEG3 (maternally expressed 3), a tumor suppressor, can downregulate miR-31 expression, which contributes to the inhibition of CRC cell proliferation and migration [81]. All these data indicate that miR-31 may serve as another significant marker of CAC and CRC, constituting a potential therapeutic target.
miR-200 familyThe miR-200 family consists of miR-200a, miR-200b, miR-200c, miR-141, and miR-429. These are the microRNAs that are significant for EMT. The role of the miR-200 family in carcinogenesis through that process refers not only to CRC, but also to other types of tumors [52]. Recent studies confirmed that the levels of miR-200 family increase in UC dysplastic lesions, while miR-200-3p is raised in CRC complications [32].
Different results were reported in a study by Peng S et al., which focused on the role of miR-200a in DSS-induced colitis. The authors performed a study on mice treated with DSS, which revealed that the level of miR-200a was overexpressed for four weeks using an adeno-associated viral (AAV) vector. Furthermore, the authors verified if an overexpression of miR-200a may influence the induction of inflammation. It turned out, that the overexpression of miR-200a resulted in the activation of the Nrf2 antioxidant pathway, thereby alleviating adverse alterations in animal and cellular models1. miR-200a was found to repress Keap1 (Kelch-like ECH-associated protein 1), a protein involved in the regulation of the Nrf2 antioxidant pathway. Given these facts, it seems miR-200a acts as a protective agent against DSS-induced colonic damage through the activation of the Keap1/Nrf2 signaling pathway [82].
The study conducted by Xiao et al. revealed that Clostridium butyricum, a human probiotic, alleviates inflammation and cancerogenesis in TNBS/AOM-induced CAC. Moreover, C. butyricum decreases cancer cell proliferation and pro-inflammatory cytokine production in cancer cell culture in a miR-200c-dependent way [83].
Olrau et al. [84] reported that miR-200a is increased in the colonic tissue of patients with UC-associated dysplasia; however, potential targets of this miRNA were not evaluated. In addition, Carter et al. proved that the increased expression of the miR-200 family in colon cancer negatively correlated with the level of the tumor suppressor gene RASSF2 (Ras association domain-containing protein 2), associated with the MAPK/ERK (mitogen-activated protein kinases/extracellular signal-regulated kinases) signaling pathway, suggesting that RASSF2 may be a direct target of miR-200 [85]. Given the discrepancies in reports on the involvement of miRNAs from the miR-200 family, further research is necessary to fully elucidate their role in CRC pathogenesis and their links to inflammation.
miR-26b familymiR-26 family, including miR-26a, miR-26b, miR-1297, and miR-4465, is associated with many types of cancer, such as gastric, lung, or prostate cancer. Depending on tumor type, their levels may be up- or downregulated and act as pro- and anticancerogenic agents [86].
Regarding CRC, data regarding the role of miR-26a are inconclusive; some reports indicate that lower levels of miR-26b were associated with neoplasm progression. Intriguingly, the study performed by Benderska et al. revealed that miR-26b expression is elevated in AOM/DSS-induced CAC. Authors suggest that miR-26b regulates the expression of proteins interacting with the DAPK (death-associated protein kinase 1) signaling pathway; especially the DIP1 protein (disco-interacting protein 1), which contains a putative miR-26b-binding domain. Furthermore, the authors reported that the level of miR-26b is higher in UC patients with neoplastic lesions compared with nonneoplastic patients and patients with the disease in remission. Authors reported that miR-26b was also elevated in celiac disease [20].
Moreover, Zhang et al. reported that transgenic mice overexpressing miR-26a are less vulnerable to AOM/DSS-induced CAC; miR-26a levels were inversely correlated with IL-6, STAT3, and NF-kB levels [87, 88]. In contrast, Fan et al. proposed that miR-26b silences the expression of PTEN and wingless-type MMTV integration site family member 5A (WNT5A), which affect EMT and tumor cell motility. This suggests that elevated miR-26b levels in CRC patients with lymphatic metastases may enhance the invasive potential of cancer cells [89].
Also interesting are the reports of Wang et al. that a higher level of miR-26b expression positively affects 5-fluorouracil (5-FU) treatment in vitro studies using CRC lines. The authors also postulate that this mechanism may be related to targeting P-glycoprotein (Pgp), which activates the intrinsic apoptotic pathway [90].
Due to these discrepancies, further research is required to clarify the role of this miRNA family in CRC development and progression.
Other miRNAs associated with CAC and CRC pathogenesismiR-19a seems to be one of the factors promoting colitis and the development of CAC through its connection with TNF-α and NF-κB signaling. miR-19a directly targets TNF-α, and the expression of TNF-α, IL-8, and granulocyte–macrophage colony-stimulating factor (GM-CSF) are significantly upregulated by a miR-19a inhibitor. In a study by Huang et al. [55], the role of miR-19a in the connection between metastatic involvements of lymph nodes and TNF-α-induced EMT in CRC was confirmed. It was also observed that the TNF-α stimulation is related to the overexpression of miR-19a in HCT-116 cells. Wang et al. found a relationship between miR-19a, NF-κB, and TNF-α signaling in CAC. They showed that TNF not only led to the overexpression of miR-19a, but the high level of miR-19a induced by TNF may be decreased through inhibition of NF-κB signaling. It was also found that the miR-19a activated the NF-κB signaling pat
Comments (0)