
Remember me

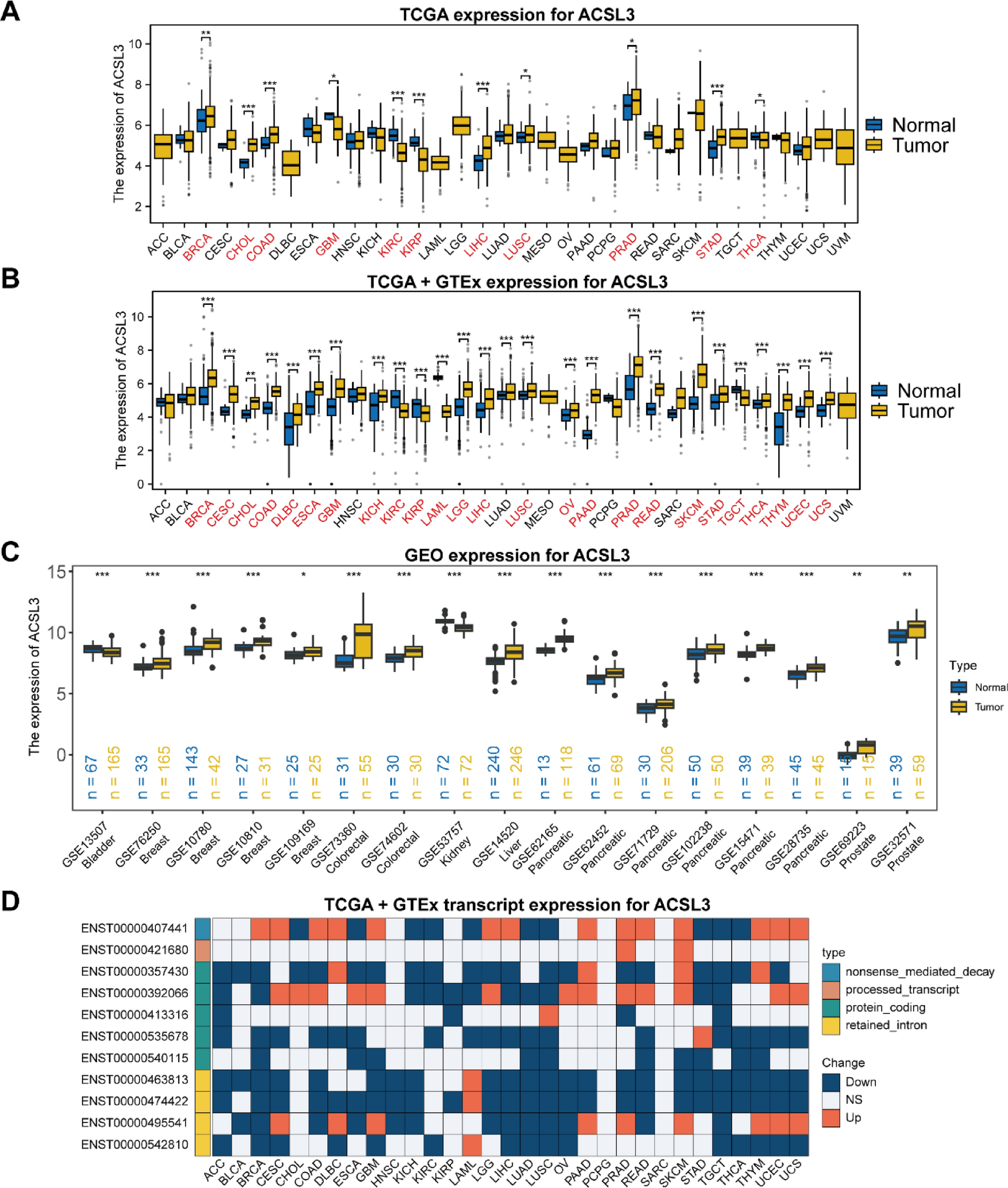
Based on TCGA and GTEx expression datasets, we analyzed ACSL3’s differential expression patterns across a range of tumors versus normal tissues. Overall, ACSL3 was found to be dysregulated in the majority of tumors, predominantly showing upregulation. Tumors with significantly higher ACSL3 expression included BRCA, CESC, CHOL, COAD, DLBC, ESCA, GBM, KICH, LGG, LIHC, LUSC, OV, PAAD, PRAD, READ, SKCM, STAD, THCA, THYM, UCEC, and UCS. Conversely, ACSL3 expression was downregulated in KIRC, KIRP, LAML, and TGCT (Fig. 1A and B). Validation of ACSL3 expression levels across 17 cancer cohorts from the GEO database yielded consistent results (Fig. 1C).
Fig. 1
Abnormal Expression of ACSL3 in various cancers. A Expression analysis of ACSL3 in the TCGA pan-cancer cohort. B Investigation of ACSL3 expression combining TCGA and GTEx data across multiple cancer types. C Examination of ACSL3 expression in the GEO pan-cancer cohort. D Transcript-level assessment of ACSL3 expression in various cancers. The statistical method used is the Wilcoxon rank-sum test. *P < 0.05, **P < 0.01, ***P < 0.001
Due to the extensive dysregulation of ACSL3 observed across cancer types, we examined its transcript levels in both normal and tumor tissues. Our analysis uncovered notable variations in transcript abundance among different cancers. Remarkably, one transcript linked to nonsense-mediated mRNA decay (NMD) (ENST00000407441) showed marked upregulation in several tumor samples, which may result from NMD evasion, gene mutations, or irregular splicing. Furthermore, the dysregulation of several non-coding ACSL3 transcripts suggests that ACSL3 might influence tumor development and progression through non-coding RNA pathways (Fig. 1D).
Expression distribution of ACSL3 in single cellsFollowing our preliminary investigation of ACSL3 levels in different cancers, we observed that the gene is broadly dysregulated, with a tendency toward higher expression in many tumors. To better understand the cell-specific distribution of ACSL3 in these cancers, we examined single-cell transcriptomic datasets. The results showed that ACSL3 is widely expressed in various immune cells and malignant clusters; however, its expression patterns are specific to different cancer cells (Fig. 2). Notably, ACSL3 has high expression levels and proportions in macrophages across all cancers, suggesting that ACSL3 may influence tumor development through dysregulated expression in macrophages, potentially reflecting the lipid metabolism reprogramming needs of tumor-associated macrophages.
Fig. 2
The expression of ACSL3 at the single-cell level across diverse cancer types. In this visualization, bubble sizes indicate the percentage of cells expressing ACSL3, while the intensity of the bubble color reflects the relative level of expression in those cells
Prognostic and diagnostic value of ACSL3 in pan-cancerWe assessed the prognostic value of ACSL3 across a range of cancers by implementing univariate Cox regression and Kaplan–Meier survival methods. These analyses examined how ACSL3 expression relates to four key survival metrics: OS, DSS, DFI, and PFI. Our findings indicate that increased ACSL3 expression is significantly associated with reduced survival in cancers such as ACC, CESC, HNSC, KICH, LAML, LGG, LIHC, LUAD, and UVM, while it appears linked to better outcomes in KIRC, OV, THCA, THYM, UCEC, and UCS (Fig. 3A). To further explore the connection between ACSL3 and clinical characteristics, we evaluated its expression in relation to patient age, TNM staging, and overall tumor stage. The analysis revealed that ACSL3 expression correlates with specific clinical features in various cancer types; for example, its levels are notably elevated in advanced stages of LIHC, LUAD, and UCS, whereas lower levels are observed in KIRC and OV (Fig. 3B). In addition, ROC analysis based on ACSL3 expression demonstrated robust diagnostic capability in several cancers, particularly in LAML, PAAD, READ, and SKCM (Fig. 3C).
Fig. 3
Prognostic and Diagnostic Value of ACSL3 in Pan-Cancer. A Analysis of ACSL3 Expression in Relation to Prognostic Outcomes (OS, DSS, DFI and PFI). B Correlation Analysis of ACSL3 Expression with Clinical Features: This part examines how ACSL3 expression relates to clinical characteristics such as age, TNM stage, and tumor stage. C ROC Analysis of ACSL3 Expression for Cancer Diagnosis: This section presents the diagnostic efficacy of ACSL3 expression across various cancers, highlighting its potential as a biomarker
Gene set enrichment analysisTo examine the biological functions of ACSL3 in cancer progression, we divided cancer samples based on ACSL3 expression levels into high and low groups. We performed differential gene expression analysis and conducted gene set enrichment analysis (GSEA). The KEGG analysis revealed that several cancers were enriched in pathways such as calcium signaling, cAMP signaling, and ECM-receptor interaction, which are upregulated and associated with cancer. Conversely, pathways like primary immunodeficiency were downregulated across multiple cancers (Fig. 4A). In the Hallmark analysis, cell cycle-related pathways, including G2M checkpoint and E2F targets, were significantly activated in numerous cancers. The activation of genome stability pathways, such as mitotic spindle, may contribute to uncontrolled tumor cell growth. Other pathways related to cancer, such as KRAS signaling, interferon alpha response, and inflammatory response, were also enriched (Fig. 4B). Interestingly, certain cancers like TGCT exhibited notable heterogeneity, with enrichment patterns differing from other cancers.
Fig. 4
Gene Set Enrichment Analysis. A KEGG GSEA Analysis Between High and Low ACSL3 Expression Tumor Patients. B Hallmark GSEA Analysis Between High and Low ACSL3 Expression Tumor Patients
Immunological characteristics of ACSL3 in the pan-cancer microenvironmentThe progression of malignant tumors is closely linked to the immune microenvironment surrounding them. To understand how ACSL3 affects immune status in different cancers, we assessed the activity scores of the anti-cancer immune cycle. Our analysis revealed that ACSL3 expression is positively associated with cancer cell antigen release (Step 1), antigen presentation (Step 2), and T cell recognition of cancer cells (Step 6). Conversely, it is negatively associated with priming and activation (Step 3), immune cell trafficking to tumors (Step 4), and immune cell infiltration into tumors (Step 5) (Fig. 5A). The ESTIMATE analysis showed that ACSL3 has a negative correlation with immune and stromal scores in several cancers, indicating that ACSL3 might influence the tumor microenvironment by affecting immune cell infiltration or stromal components (Fig. 5B). Additionally, analysis of immune cell infiltration levels across cancers indicated that, apart from T helper cells, Tcm, Tem, and Th2 cells, most immune cells exhibit a negative correlation with ACSL3 expression (Fig. 5C).
Fig. 5
Immunological characteristics of ACSL3 in pan-cancer. A Variations in anticancer immune cycle activity scores are observed between tumor patients with elevated and reduced ACSL3 expression levels. B ESTIMATE analysis assessing the correlation between the tumor immune microenvironment and ACSL3. C ssGSEA analysis evaluating the relationship between tumor immune cell infiltration and ACSL3
Regulatory factors related to the immune system are essential in shaping the tumor microenvironment and affecting the success of cancer immunotherapy. Therefore, we also studied the association between ACSL3 and MHC molecules and immune checkpoints. The results showed significant associations, although the correlations varied among different cancers and molecules. Among MHC molecules, ACSL3 is positively correlated with TAPBP, TAP2, TAP1, and B2M in most cancers, while it is largely negatively correlated with other molecules (Fig. 6A). Immune checkpoints generally showed a positive correlation with ACSL3 across most cancers, while TMIGD2, TNFRSF14, TNFRSF18, and TNFRSF25 were positively correlated with ACSL3 (Fig. 6B). Overall, cancer patients with high ACSL3 expression exhibit inactive immune cell activity, presenting an immunosuppressive state.
Fig. 6
Correlation of ACSL3 with immune-related molecules in pan-cancer was investigated. A We analyzed the relationship between ACSL3 and MHC. B We examined the association between ACSL3 and immune checkpoints
Genetic alteration analysis of ACSL3 in pan-cancerGenomic stability influences gene expression and tumor progression, and is linked to immunotherapy responses. Analyses of TMB and MSI showed a positive correlation between ACSL3 expression and TMB scores in cancers like SARC, PAAD, THYM, STAD, SKCM, GBM, UCEC, COAD, OV, LUSC, and LUAD. Similarly, ACSL3 expression positively correlates with MSI scores in cancers such as READ, MESO, UCEC, KIRC, COAD, SARC, LUSC, and STAD. In contrast, ACSL3 expression is negatively correlated with TMB scores in THCA, PRAD, KIRC, and KIRP, and with MSI scores in BRCA, THCA, PRAD, and DLBC (Fig. 7A and B). These results indicate that ACSL3 might be a predictive biomarker for cancer immunotherapy effectiveness in certain cancers.
Fig. 7
Analysis of ACSL3 genetic alterations in pan-cancer. A examining the relationship between ACSL3 and tumor mutational burden (TMB). B assessing the link between ACSL3 and microsatellite instability (MSI). C evaluating copy number variations in the ACSL3 gene. D exploring the connection between ACSL3 and DNA methylation sites. E investigating correlations between ACSL3 and RNA modification enzymes, including those related to m1A, m5C, and m6A
To explore the mechanisms underlying ACSL3 dysregulation in pan-cancer, we analyzed the genetic alterations of ACSL3. We observed diverse types and frequencies of genetic alterations in ACSL3 across different cancers. Most cancers exhibited ACSL3 mutations, especially UCEC, while copy number amplification predominated in PAAD and UCS (Fig. 7C). ACSL3 expression showed a significant correlation with DNA methylation sites across various cancers, suggesting that DNA methylation influences ACSL3 regulation (Fig. 7D). Notably, compared to promoter regions, more methylation sites in gene body regions were associated with ACSL3 expression. Generally, promoter region methylation silences gene expression, while gene body methylation is sometimes linked to gene activation and transcription maintenance, although further research is needed to clarify these findings. Overall, these results highlight the complex regulatory mechanisms of ACSL3 expression.
RNA modifications, as key post-transcriptional regulatory mechanisms, play critical roles in gene expression regulation. Dysregulation of RNA modification enzymes has been shown to be closely associated with tumor initiation, progression, and metastasis. Our pan-cancer analysis revealed that ACSL3 expression levels were significantly positively correlated with various RNA modification enzymes, including those involved in m1A, m5C, and m6A modifications (Fig. 7E). This finding suggests that ACSL3 expression may be profoundly influenced by an aberrantly active RNA modification regulatory network.
Analysis of potential therapeutic drugsTo explore therapeutic agents targeting ACSL3-related tumor-promoting pathways, we analyzed the top 100 most significantly differentially expressed genes for each cancer type from a pan-cancer differential analysis. These genes underwent correlation analysis with gene expression data from the CMAP database. This approach identified the top 30 compounds with negative correlations, such as GW-8510, lanatoside_C, and 6-azathymine, which may potentially counteract cancer-associated gene expression patterns (Fig. 8A).
Fig. 8
Exploration of Potential Therapeutic Agents. A Identified 30 compounds from the CMAP database showing negative correlation with ACSL3 expression. B Conducted drug sensitivity analysis using data from the CellMiner database
Additionally, we examined FDA-approved and clinically validated drugs from the CellMiner database. Six anti-tumor drugs were found to have expression sensitivity correlations with ACSL3. Notably, HYPOTHEMYCIN exhibited a positive correlation, indicating that patients with elevated ACSL3 expression might respond better to HYPOTHEMYCIN treatment (Fig. 8B).
Knockdown of ACSL3 inhibits proliferation and migration of liver cancer cellsTo enhance the reliability of our research, we conducted experimental validation in liver cancer. First, we verified the significant upregulation of ACSL3 expression in HCC across 13 independent cohorts (Fig. 9A). We explored the expression levels of ACSL3 in various liver cancer cell lines using the Cancer Cell Line Encyclopedia (CCLE) database, and found that ACSL3 has higher expression levels in Huh-7 and HepG2 cell lines (Supplementary Table 2). Subsequently, we constructed ACSL3 knockdown cells in liver cancer cell lines Huh-7 and HepG2. The knockdown efficiency was validated using real-time quantitative PCR and Western blot (Fig. 9B and C). Based on the knockdown efficiency, we selected Huh-7 cells for further experiments. Colony formation assays showed that knockdown of ACSL3 reduced the number and size of colonies formed by Huh-7 cells, indicating weakened long-term proliferation ability (Fig. 9D). Transwell assays demonstrated that knockdown of ACSL3 decreased the migration and invasion capabilities of Huh-7 cells (Fig. 9E). CCK8 assays also indicated that knockdown of ACSL3 reduced the proliferation ability of Huh-7 cells (Fig. 9F). Wound healing assays confirmed that ACSL3 silencing significantly reduced the wound healing rate at designated time points, suggesting impaired migration (Fig. 9G). Finally, AV/PI staining analysis showed that inhibiting ACSL3 expression significantly promoted apoptosis in Huh-7 cells (Fig. 9H).
Fig. 9
In Vitro Studies on ACSL3 Knockdown and Its Effect on Liver Cancer Cell Growth. A Confirmed ACSL3 overexpression in liver cancer across 13 separate cohorts. B, C Huh-7 and HepG2 cells were transfected with shRNA to target ACSL3, and knockdown efficiency was measured at mRNA and protein levels using real-time PCR and Western blotting. D Colony formation tests. E Transwell migration assays. F Cell proliferation evaluated with Cell Counting Kit-8. G Wound healing assays assessed cell migration. H Annexin V-FITC/PI staining used in flow cytometry analysis
To further verify the role of ACSL3 in tumor cell proliferation, we injected control or ACSL3 knockdown Huh-7 cells into BALB/c nude mice (Fig. 10A). The results indicated that knockdown of ACSL3 significantly reduced tumor growth rate and volume (Fig. 10B and D).
Fig. 10
In Vivo Experiments on ACSL3 Knockdown Inhibiting Proliferation of Liver Cancer Cells. A Schematic diagram of the in vivo experimental design. B Tumor growth curves in vivo. Tumor volume was measured at different time points. C Body weight changes in mice. D Tumor weight and morphology at the endpoint
Comments (0)