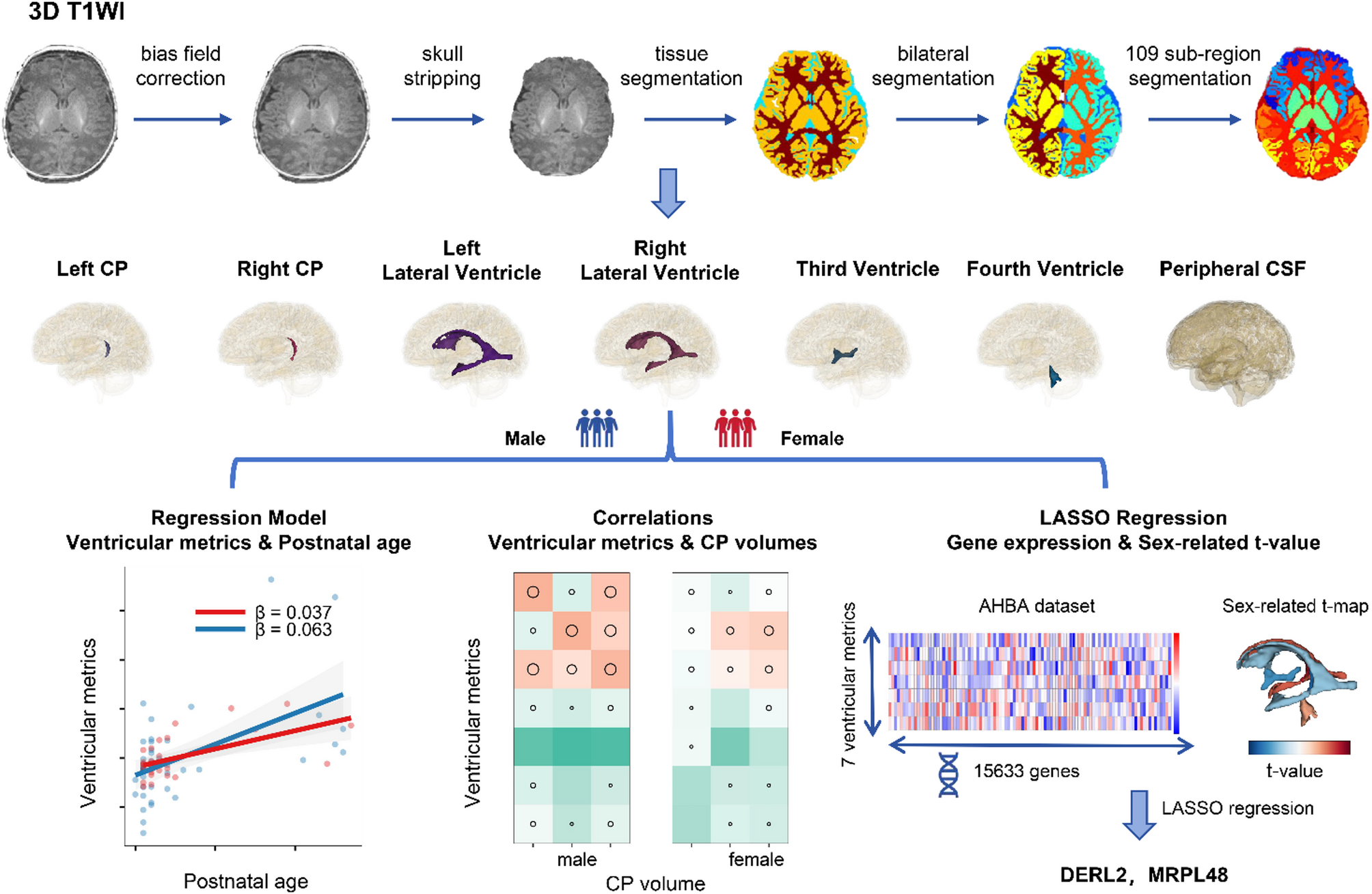
This study of 75 full-term healthy neonates utilized deep-learning automated segmentation to analyze metrics of the brain ventricular system and CSF. It was found that males exhibit larger ventricular CSF spaces and faster growth rates than do females. Even after adjusting for TBV, males continued to demonstrate a positive linear growth trend, with females showing a relatively stable pattern. We hypothesize that these differences are partly due to variations in the volume and growth patterns of the CP between sexes.
CP as a source of CSF differences
The CP, which is the primary source of CSF, is essential for the secretion and transport of CSF and for maintaining the blood–CSF barrier [28]. During embryo development, the CP first appears in the fourth ventricle, followed by the simultaneous formation of the CP in the lateral ventricles, and lastly in the third ventricle [5]. CP epithelial cells have been implicated in the pathophysiology of various neurological conditions, including neurodegenerative diseases, hydrocephalus, and stroke [29]. While MRI studies have predominantly focused on the role of the CP in neuroinflammation in conditions such as multiple sclerosis, where an increased CP volume is correlated with greater disability and cognitive decline [30], its growth trajectory and sex-related differences at the macrostructural level in neonates have been largely overlooked. The CP was segmented in our study primarily into the lateral ventricles due to the limited visibility and impact of that in third or fourth ventricles [31]. Our study suggests that sex-related differences in CP volume and growth patterns are a key factor contributing to the observed variations in the CSF system between male and female neonates. Similar sex-related differences were found both in right lateral ventricles and right CP volume. There was also a stronger correlation between right CP and right lateral ventricle volumes in males than in females. Moreover, the growth pattern of the CP closely aligns with those of the corresponding lateral ventricles, third ventricle, peripheral CSF, and the total CSF volume, indicating a possible mechanistic link. Previous research has shown that CP epithelial cells contribute to neurodevelopment and cognitive function [6]. which underscores the potential impact of CP differences on CSF dynamics and brain development.
Larger volume and faster growth of lateral ventricles in males
Our study has confirmed that males have larger lateral ventricles with faster growth rates, especially for the right lateral ventricle. The lateral ventricles in fetuses were previously reported to be 22.1% larger in males [9]. A study focusing on white matter and subcortical gray matter, and which did not consider the CSF volume, found that 1-month-old infants underwent age-related changes, with males showing a larger TBV and region-specific differences after correcting for TBV [12]. Research involving full-term or term-equivalent preterm infants aged 2 to 90 days has identified lateral-ventricle asymmetry and a significant interaction between sex and age, although the CP was not examined [32]. Additionally, a study of infants aged 3–13 months found that males had larger lateral ventricles than females, with this sex-related difference disappearing when the volumes were normalized [33]. This is consistent with findings in studies of children aged 0–11 years [34] and 0–18 years [10]. While these studies have consistently shown that males have a larger ventricular system, this sex-related difference is somewhat diminished when TBV is accounted for. Our findings suggest that sex-related differences in the ventricular system and CSF also exist during the neonatal period, aligning with those observed at other ages and in other brain regions during development. While the left lateral ventricle is generally larger than the right lateral ventricle, our findings indicate that the key sex-related difference in the lateral ventricles is on the right side, which aligns with the difference in right CP volume. This suggests that factors beyond TBV, such as sex-related brain lateralization, contribute to these differences. Notably, significant brain asymmetries and sex-related differences have been observed early in childhood [35], with structures such as the globus pallidus and putamen that are part of the ventricular wall [36] and influence cognitive function.
Regarding development rates, previous studies have shown that the absolute volume of CSF increases significantly during the second half of the gestation period, while the volume ratio of CSF to brain parenchyma stabilizes over time [9]. The ventricle volume was reported to increase by 350% from a GA of 15 weeks to birth, and by 86% from birth to 1.25 years, while it decreased by 18% from 1.25 to 3.5 years [34]. CSF volume hasn’t reached the peak during the pediatric age range [10]. Our findings, consistent with the stabilizing yet increasing growth of CSF space from gestation to birth, have revealed the significant sex-related differences in the early postnatal growth patterns of the lateral ventricles after adjusting for GA. This information might be particularly useful when developing optimal protocols for monitoring sex-related development during the neonatal period.
However, it is crucial to interpret these findings with caution. In the cohort, the majority of MRI scans were acquired within the first week after birth, resulting in a highly skewed distribution of postnatal age. Although significant associations were detected between postnatal age and CSF-related metrics, these likely reflect developmental dynamics specific to the immediate postnatal period, rather than the full neonatal stage. Therefore, our results should be viewed as preliminary and time-specific. Future research incorporating more temporally distributed data across the neonatal period—and ideally longitudinal designs—will be essential to better characterize the evolving trajectory of CSF space development and its sex-specific patterns.
No difference in total CSF volume with relatively faster growth of peripheral CSF in males
Our study suggests that peripheral CSF constitutes a substantial portion of total CSF, likely mitigating sex-related differences in lateral ventricles, leading to the absence of differences in total CSF volume. The growth pattern of peripheral CSF aligns with the sex-related differences observed for other CSF space metrics: while males continue to show growth, the values for females stabilize. Our findings reveal significant sex-related differences in CSF distribution patterns and growth rates. These differences may be influenced by biochemical factors such as the osmolarity, pressure, and flow rate of CSF [6]. Previous research studies have found that males generally have higher CSF protein content and albumin quotient values across various pathologies and age groups, and are more likely to experience blood–CSF barrier dysfunction [2].
Other findings in third and fourth ventricles, and in EI
The third ventricle showed significant growth in both its absolute volume and its percentage relative to TBV. Previous studies indicate that the GA at birth does not significantly influence the size of the third ventricle during the first 3 months of life [32]. However, a study involving 700 children found that during normal development the third ventricle exhibited a segmental increase from birth to the age of 18 years, with no influence of sex [37]. Our findings suggest that the third ventricle exhibits a delayed growth phase during fetal development, followed by rapid expansion during the neonatal period. Positioned between the two halves of the thalamus, the third ventricle is essential for CSF circulation [38] and participates in the contributions of the thalamus to the regulation of vital functions [38, 39] and to the development of higher brain functions [40].
In contrast, the percentage of the fourth ventricle relative to TBV exhibited a declining trend in the present study. Since the fourth ventricle serves as a critical junction between the ventricular system and the subarachnoid space, its obstruction can lead to rapid CSF accumulation that represents a medical emergency [41]. The early detection of subtle changes in the fourth ventricle may therefore have important clinical implications for the prompt diagnosis and management of neurodevelopmental disorders.
The EI is commonly used in clinical practice as a rapid alternative to MRI volumetric quantification for conditions such as pediatric hydrocephalus [42], normal-pressure hydrocephalus [43], and Alzheimer’s disease (AD) [44]. The present study has revealed significant sex-related differences in the EI, suggesting that including sex-related differences when calculating the EI will improve clinical diagnoses.
This study was subject to several limitations that should be addressed in future research. First, the smallness of the sample reduces the generalizability of our findings. Increasing the sample size in future studies will enhance the robustness of the results and allow for more-nuanced analyses of sex-related differences in the ventricular system and CSF development. Second, the autosegmentation of small and intricate structures such as the third ventricle, fourth ventricle, and CP may not be as precise as metrics involving structures such as white matter, which potentially obscures subtle differences. Employing more-advanced imaging techniques or manual segmentation could improve accuracy. Third, our study did not account for socioeconomic status or maternal health during pregnancy, and these factors could influence neonatal brain development. Future research should include matched cohorts to control for these variables. Finally, this was a single-center study, which may reduce the external validity of our findings. Multicenter longitudinal studies are needed to confirm these results and further explore the sex-related differences in CSF space metrics.
Insights from gene expression
Using Lasso regression to predict sex-related differences in the left and right CPs, lateral ventricles, third and fourth ventricles, and peripheral ventricles based on regional gene expression, we identified two relatively important genes: DERL2 and MRPL48.
DERL2 is a key component of the endoplasmic reticulum quality control system and plays a essential role in the degradation of misfolded glycoproteins through the ER-associated degradation (ERAD) pathway [45, 46]. Maintaining ER homeostasis is critical for normal brain development and memory formation [47], whereas disruption of this balance has been implicated in neurodegenerative diseases characterized by the accumulation of misfolded proteins [47]. In AD model mice, physical exercise has been shown to upregulate DERL2 expression in the hippocampus [48]. Global knockout of DERL2 results in perinatal lethality, and the few surviving mice exhibit skeletal dysplasia due to defective collagen secretion in costal chondrocytes [49]. Conditional knockout of DERL2 in the CNS impairs postnatal brain development, particularly affecting the cerebellum and striatum, and leads to motor deficits [50]. DERL2 deficiencies has also been shown to inhibit neurite outgrowth and suppress sterol regulatory element-binding protein 2-mediated brain cholesterol biosynthesis—an essential process for glial proliferation, neurite extension, and microtubule stability [51]. In parallel, CSF system also contributes significantly to the clearance of misfolded proteins. Impaired meningeal lymphatic drainage, especially during aging or in AD, can exacerbate toxic protein accumulation in CNS [52].
MRPL48 is a component of the 39 S large subunit of the mammalian mitochondrial ribosome and plays a critical role in mitochondrial translation [53]. Encoded by nuclear DNA, MRPL48 exhibits region-specific expression in the brain, with significantly higher levels observed in the frontal cortex compared to the hippocampus or hypothalamus in rats [54]. In addition, MRPL48 has been implicated in the development of nervous and endocrine system, with its dysregulation linked to hereditary, metabolic, and neurological disorders [55]. In forebrain mineralocorticoid receptor knockout mice, MRPL48 expression is downregulated, while glucocorticoid receptor expression is slightly upregulated [56]. Notably, MRPL48 displays a female-specific paternal expression bias in the medial preoptic area [57] and is differentially expressed in patients with schizophrenia compared to healthy controls [58]. Moreover, a study using cDNA microarray and proteomic analyses in rats have shown that gene expression in the choroid plexus is enriched in pathways related to mitochondrial dysfunction and oxidative phosphorylation [59].
In conclusion, the downregulation of DERL2 and upregulation of MRPL48 observed in our study may contribute to the altered clearance of misfolded proteins and altered energy metabolism.These molecular alterations could ultimately underlie sex-related morphological differences in the ventricular system (Fig. 6).







Comments (0)