John N. Stallone, PhD, FAPSIntroduction
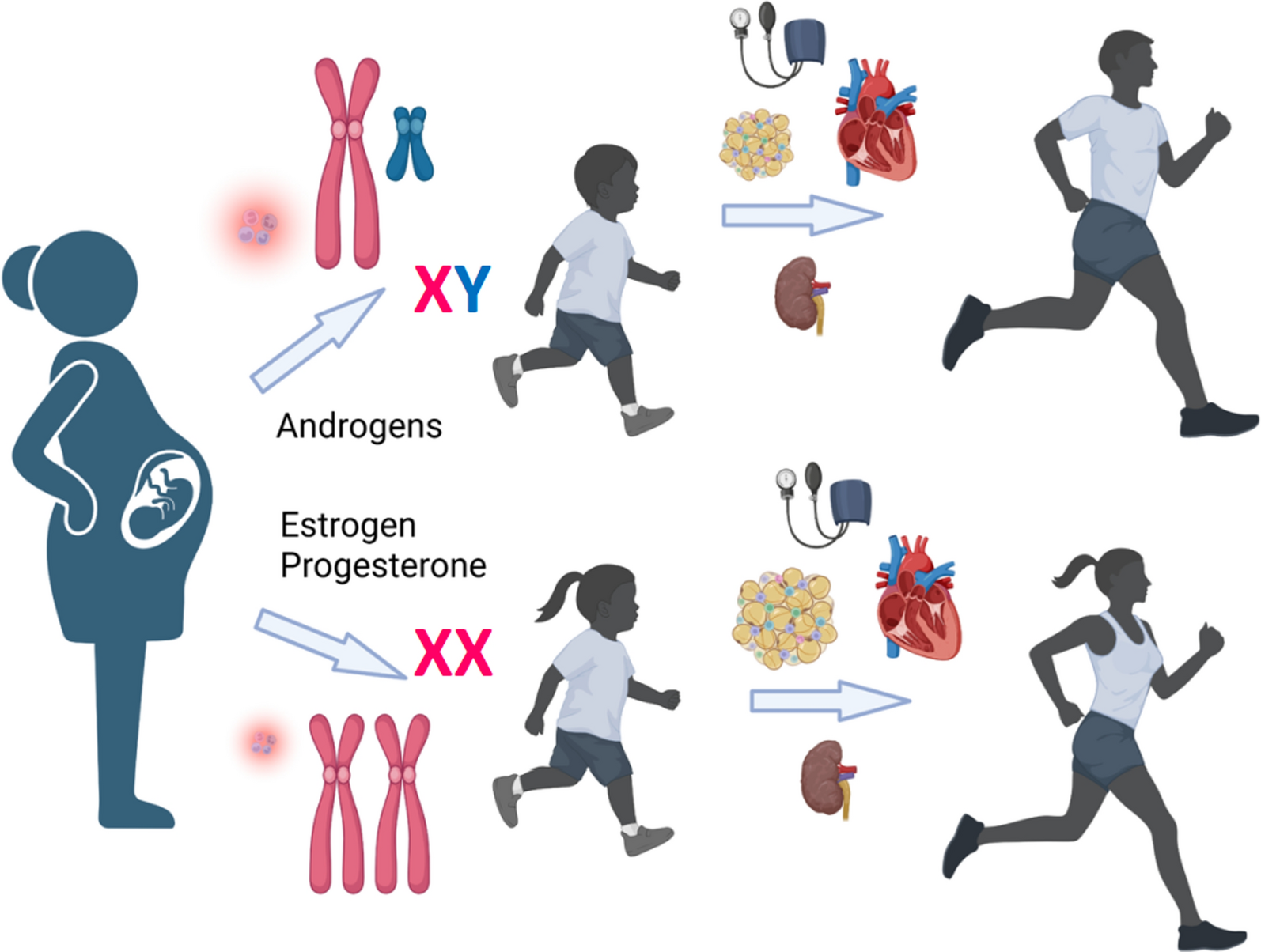
The male sex steroid hormones (testosterone (TES) and related C19 androgen molecules) are the humoral messengers responsible for the differentiation, development, and maintenance of the male reproductive system and secondary sexual characteristics that differ so markedly between males and females. Although the anatomical, behavioral, and reproductive effects of TES were recognized in animal experiments as early as the eighteenth and nineteenth century (long before its chemical structure as the principal mammalian male sex hormone of testicular origin was identified in 1935) [175, 176], it is now clearly recognized that the androgens exert a broad variety of physiologically relevant effects on the regulation of cardiovascular, hematopoietic, immune, metabolic, and nervous systems as well as their well-known anabolic effects on bone and skeletal muscle [177, 178]. It is important to recognize that the androgens are synthesized in both sexes and circulate in the plasma, albeit at quite different levels. Plasma levels of TES in females are 5–10% of those in males; however it is increasingly recognized that the androgens play unique roles in the regulation of reproductive as well as cardiovascular, metabolic, and other functions in females as well as males [177, 178].
Sex differences in the cardiovascular effects of androgens
Cardiovascular diseases (CVD) are a major cause of morbidity and mortality and in the Western world, one-third of all deaths are attributable to CVD [179]. A conspicuous feature of this healthcare epidemic is that most forms of CVD are higher in men than in premenopausal women, yet the reasons for these prominent sex differences remain unclear. The clinical case studies and epidemiological observations that hypertension (HT) and coronary artery disease (CAD) occur more frequently in men than in premenopausal women [180,181,182,183,184,185,186] have led to the dogmatic view that TES and other androgens exert deleterious effects on the heart and vasculature and worsen the development of CVD in men, in part by exacerbating risk factors such as blood pressure (BP), body fat composition, insulin resistance, and serum lipid profiles [187,188,189]. In parallel, earlier animal studies provided support for this dogma and revealed that in various rat models of HT castration attenuates the development of HT in males [190,191,192,193,194]. However, more recent human clinical trials [195,196,197] and experimental animal studies [198,199,200,201] reveal that TES and its metabolites exert beneficial effects on BP and metabolic function in males, which are risk factors for CVD. Reconciliation of the conflict between earlier and more recent studies on the cardiovascular effects of TES depends upon careful scrutiny of earlier experimental animal and human studies which suffered from flaws or limitations in experimental design and/or the animal models employed [184, 186, 196]. For example, most earlier animal studies employed unrealistic models and short term settings. Similarly, subsequent analyses of earlier human clinical and epidemiological studies revealed important validity issues with experimental design, data collection and analysis, and selective exclusion of data, which emphasize the importance of careful study design and that dogma and controversy can adversely distort the validity of human clinical and epidemiological findings concerning TES (for detailed review of these issues, see [202]).
Human clinical and experimental animal studies have clearly established that TES and other androgen metabolites exert beneficial effects by inducing relaxation of vascular smooth muscle (VSM) through rapid, nongenomic (androgen receptor (AR)-independent) mechanisms in vitro (for recent reviews, see [203,204,205]. Although this acute effect of TES and other androgens was initially reported at high (micromolar) concentrations in variety of large arteries from several species, more recent studies revealed that TES produced relaxation of smaller resistance arteries and arterioles at nanomolar (physiological) concentrations (i.e., mesenteric, prostatic, pulmonary, and subcutaneous) (for reviews, see [202,203,204,205]. The key mechanism underlying this effect of TES on VSM appears to be activation of calcium-dependent (BKCa) and voltage-operated (KV) K+ channels via TES-induced activation of neuronal nitric oxide synthase (nNOS) and/or inactivation of L-type voltage-operated Ca2+ channels in VSM [204, 205]. Although numerous studies have clearly established that TES and other androgens exert rapid, nongenomic vasorelaxing effects in vitro, evidence that TES produces coronary or systemic vasodilation in vivo at physiological concentrations (100 pM to 100 nM) is more limited. Studies in anesthetized dogs [206], pigs [207] and humans [208] demonstrated that intra-arterial infusions of TES produces coronary vasodilation, and regional vasodilation of mesenteric, renal, and skeletal muscle vascular beds in anesthetized pigs [207]. More recently, several studies reported that TES and other androgens produced systemic vasodilation. In conscious, ganglionic-blocked male Sprague–Dawley (SD) rats, i.v. infusion of TES or its genomically inactive metabolite 5β-dihydrotestosterone (5β-DHT) produced dose-dependent systemic hypotension [198]. Similarly, bolus i.v. injections of TES, 5β-DHT, and the potent genomically active metabolite 5α-DHT produced dose-dependent hypotension in Spontaneously Hypertensive (SHR) and to a lesser extent in normotensive-control WKY male rats [199]. In both studies, the hypotensive effect of 5β-DHT was more efficacious than that of TES. While these recent studies clearly reveal that exogenous TES and its metabolites exert important hypotensive effects on systemic BP through direct vasodilatory actions on the systemic vasculature, the role of endogenous androgens in the long-term regulation of BP remained unanswered until recently. Long-term studies by Perusquia et al. [200] revealed that castration of both Wistar and WKY male rats led to the progressive development of HT over a period of 11 weeks that then plateaued through 18 weeks (151 ± 2 vs. 110 ± 2 mmHg at baseline, mean arterial BP). Subsequent long-term studies by Hanson et al. [201]. demonstrated that castration of male SD rats produced progressive HT from 109 ± 3 at baseline to 143 ± 3 mmHg systolic BP at 10 weeks, and that subsequent TES replacement therapy to physiological levels completely normalized BP in 5 weeks to 113 ± 1.3 mmHg. Interestingly, nearly identical effects of castration and TES replacement were observed in AR-deficient Testicular-feminized male (Tfm) rats, strongly suggesting that the cardiovascular effects of TES are nongenomic in nature. Treatment of SD rats with the type 1 angiotensin receptor antagonist Losartan completely prevented development of HT. rt-PCR of the kidney revealed that castration increased expression of mRNA for renin (92%), angiotensin converting enzyme (58%), and angiotensin type 1 receptor (80%), while TES replacement therapy completely normalized renin-angiotensin system (RAS) mRNA expression to levels of intact control male rats. These findings reveal that both endogenous and exogenous TES exert anti-hypertensive effects that appear to involve reductions in RAS expression in the kidney, enhanced fluid excretion, and enhanced systemic vasodilation.
The overwhelming majority of studies on the cardiovascular effects of the androgens in experimental animals, and to a lesser extent in human clinical trials, have employed males. However, given that measurable levels of TES and other androgens are present in the circulation of female animals and humans, then it is also important that the effects of these hormones be studied in females. Studies of the acute effects of TES on blood vessels isolated from females, while limited in number, have uniformly revealed that TES-induces vasorelaxation of rat aorta, human pulmonary artery and vein, and isolated, perfused human lung do not differ between males and females [209,210,211]. Similarly, acute intra-arterial infusions of TES in anesthetized pigs produced similar regional vasodilation in females as in males [207]. In near-term normal-pregnant and preeclamptic-pregnant female Wistar rats, bolus i.v. injections of TES, 5α-DHT, 5β-DHT, and dehydroepiandrosterone (DHEA) produced similar substantial reductions in mean arterial BP in both groups, while 5β-DHT and DHEA exhibited significantly greater hypotensive potency than TES or 5α-DHT [212]. Isolated thoracic aortae from these same pregnant groups also exhibited similar vasorelaxing responses to these same androgens. With regard to long-term effects of TES treatment on vascular function and BP, several studies have examined effects in female animals and humans. Long-term treatment of female Cynomologous monkeys with TES while fed a high fat diet increased coronary arterial atherosclerotic plaques but improved vasomotor responses to acetylcholine [213]. In contrast, long-term treatment of ovariectomized female SHR rats with TES for 5–10 weeks increased arterial BP to levels similar to those of male SHR [190, 214]. Although not a goal of this study, this experimental design served as a model to test the effects of cross-sex hormone replacement in female-to-male transexuals, and resulted in similar detrimental effects as long-term TES treatment in human female-to-male transexuals, which impaired flow-mediated vasodilation [215]. Similarly, several prospective human clinical trials identified positive relationships between plasma free (bioavailable) TES levels and incidences of elevated BP, HT, and CAD in pre- and post-menopausal women [216,217,218]. While these human studies appear to conflict with the majority of experimental animal studies on the vascular effects of TES, the former studies could not discern whether elevations in plasma TES followed or preceded the increases in BP, HT, and CAD in human females. Likewise, long-term treatment of females with doses of TES that elevated plasma levels to those of males likely produce very different cardiovascular effects than the much lower normal plasma levels of TES in females. Indeed this possibility in quite likely since male levels of TES in female animals are associated with detrimental metabolic effects, while the same levels in males produce beneficial metabolic effects (for detailed review of these issues, see [202]). In summary then, while acute vascular effects of TES and other androgens are clearly beneficial and similar in males and females, long-term systemic effects of the androgens may differ in males and females and are likely related to the pronounced sex differences in plasma levels of TES, as well as the distribution and numbers of receptors present in the target tissues.
Sex differences in the metabolic effects of androgens
The early eighteenth and nineteenth century experiments of Hunter and Berthold on roosters [175, 176] and those of Brown-Sequard involving self-injection of aqueous extracts of dog and guinea pig testes (so-called organotherapy) [219] led to the recognition that a humoral substance from the testes (identified much later as TES in 1935) was responsible for beneficial effects not only on reproductive tract function, but also on overall vitality, physical strength, and intellectual capacity. As discussed above, earlier human epidemiological studies and clinical trials led to two key areas of study in the 1960s that continue today, the effects of androgens on cardiovascular and metabolic health and disease. An extensive review of these earlier studies led Kalin and Zumoff [180] to conclude that androgens exerted direct detrimental effects on the atherogenic process in the coronary arteries, contributing to the development of CAD. This view reflects the long-known major influence of androgens on body fat composition, muscle mass, and bone density in the male [220, 221] and the idea that these hormones exerted detrimental effects on lipid metabolism (e.g., dyslipidemia), an important risk factor for CVD. This belief was furthered by clinical observations that the use of anabolic steroids by athletes (synthetic derivatives of TES) to enhance muscular strength resulted in premature, higher incidences of HT, ventricular remodeling, and sudden cardiac death [222,223,224]. However, more recent human epidemiological studies and clinical trials have challenged this dogmatic view and revealed that in addition to the well-known classical effects of TES on bone density and muscle mass, that it also plays a key beneficial role in the regulation of carbohydrate, fat, and protein metabolism and inflammation, and that TES deficiency is strongly associated with increased fat mass, hyperlipidemia, hypercholesterolemia, hyperglycemia, and insulin resistance, as well as elevated BP, the cluster of symptoms comprising metabolic syndrome (MetS) and type 2 diabetes mellitus (T2DM). MetS and T2DM share a common etiology (central adiposity) and are central risk factors for CVD (for reviews see [202, 220, 221, 225]). Indeed, the link between hypogonadism and the development of visceral obesity, insulin resistance, and MetS in men has been well established by many recent studies (for review, see [226]).
Human clinical and experimental animal studies have identified several key actions of TES in the male that promote glucose and lipid homeostasis, including: prevention of visceral fat accumulation (adipogenesis), improved insulin sensitivity in adipose, skeletal muscle, liver, and brain, central (hypothalamic) effects to enhance energy expenditure and leptin sensitivity, and regulation of pancreatic β-cell function to improve glucose tolerance and glucose-stimulated insulin secretion. These findings have led to a proposed mechanism of androgen actions to promote glucose and energy homeostasis via AR-mediated effects on adipose tissue, liver, pancreatic β-cells, skeletal muscle, and metabolic centers in the hypothalamus [226]. Interestingly, aromatization of TES to 17β-estradiol that interacts with the estrogen receptor (ER) appears to be at least partly responsible for preventing abdominal obesity; thus, both TES and estrogen appear to play important roles in the regulation of energy homeostasis in males.
The dramatic sexual dimorphism in the metabolic effects of androgens in men vs. women is reflected by the striking differences in body fat distribution and skeletal muscle mass observed between the sexes, which begin at birth and are enhanced greatly at puberty with the surges of sex steroid hormone secretion. Thus, men tend to have less total body fat but more abdominal adipose tissue (i.e., android distribution) and greater skeletal muscle mass, driven by the effects of TES. In contrast, women tend to have more total body fat distributed with a gluteal/femoral and subcutaneous (i.e., gynoid) distribution and less skeletal muscle mass, driven by the metabolic actions of estrogen [227,228,229]. While estrogen in the female mediates the amount, deposition, and function of the metabolically safer gynoid body fat distribution, TES on the other hand drives the deposition and function of the more unfavorable abdominal adipose tissue in the male; however, this is metabolically compensated for by the anabolic effects of TES to increase lean tissue and skeletal muscle mass and function to impair adipogenesis.
While the role of TES in the regulation of metabolism in men and the impact of TES deficiency on the development of visceral obesity, insulin resistance, and the MetS is well established, the the role of androgens in the regulation of metabolism in females and the possible impact of abnormalities in TES levels on metabolic dysfunction in women has not been well studied [230], even though associations between androgen excess and diabetes, obesity, and infertility have been known for nearly a century. Indeed, the relationship between androgen excess and diabetes has been known since the report of “diabetes in bearded women” by Achard and Thiers in 1921 [231]. Similarly, the link between obesity and the clinical triad of polycystic ovaries, hirsutism, and oligo/amenorrhea was first reported in 1935 as the Stein-Leventhal Syndrom [232], which was later renamed as polycystic ovary syndrome (PCOS). Thus, exactly opposite of the situation in males, the coexistence of excess androgen levels with cardiovascular risk factors (i.e., dyslipidemia, insulin resistance, and obesity) and increased atherosclerosis [233] that occur in PCOS has advanced the concept that excess androgens exert adverse effects in women [234, 235].
Likewise, other hyperandrogenic conditions in women such as congenital adrenal hyperplasia (CAH) and androgenized female-to-male transexuals are also associated with metabolic dysfunction, especially glucose intolerance, insulin resistance, obesity, and subsequently with T2DM [226]. Thus, the much lower levels of TES in normal women appear to exert beneficial effects, while the elevated androgen levels in pathophysiological states such as PCOS result in cardiovascular and metabolic dysfunction; however, this sexual dimorphism in the metabolic effects of androgens in males vs. females is puzzling and poorly understood [236, 237]. The aforementioned observations have led to the concept that a delicate equilibrium exists between androgen effects on adipose tissue vs. skeletal muscle that underpins the metabolic phenotype seen with androgen excess in females vs. androgen deficiency in males. This concept of overlapping adverse metabolic effects of androgen excess in women vs. androgen deficiency in men has been termed the “metabolic valley of death” [236] (for more detailed review, see [202]).
The dramatic sexual dimorphism in the relationship between TES and metabolic function and the delicate balance of androgen effects in adipose vs. skeletal muscle raises the question of what mechanism(s) underlie the disruption of the equilibrium in TES effects? Three major factors appear to be involved; namely, gonadal dysfunction, normal aging, and sedentary lifestyle. Thus, in the male, these disturbances lead to a deficiency of testicular androgen secretion, resulting in increased abdominal fat deposition, dyslipidemia, loss of skeletal muscle mass, increased insulin resistance, and development of MetS and T2DM. Conversely, in the female, age-dependent reductions in estrogen and relatively small increases in circulating androgens from PCOS or CAH (or larger increases with cross-sex hormone therapy in female-to-male transexuals) lead not only to phenotypic masculinization, but also cause “masculinization” of adipose tissue and its conversion from gluteal/subcutaneous fat to abdominal (visceral) fat deposition with the expression of pro-inflammatory cytokines similar that observed in males [238, 239]. It has been proposed that the excess TES and AR activation lead to deleterious effects on glucose, fat, and energy homeostasis, including: activation of adipose tissue (increased adiposity and inflammation), central (hypothalamic) effects to reduce energy expenditure and leptin sensitivity, activation of macrophages (oxidative stress), excess pancreatic β-cell function (insulin secretion), and skeletal muscle insensitivity to insulin. Together, these effects synergize to promote metabolic dysfunction, inflammation, visceral adiposity, and eventually, T2DM [226].
In summary then, the available data from both experimental animal and human studies reveal the existence of a bi-directional modulation of glucose and fat homeostasis in females vs. males. Thus, androgen or AR deficiency results in dramatic metabolic dysfunction in aging males, but to a much lesser extent in females. Since AR activation is weaker in females due to (normally) substantially lower circulating androgen levels and a much smaller population of AR in metabolic target tissues, TES and other androgens are less important in the maintenance of energy homeostasis in females under normal conditions; however, the elevated levels of androgens that occur with PCOS, CAH, or other pathological conditions are sufficient to result in metabolic dysfunction. Likewise, the substantial levels of TES resulting from cross-sex hormone therapy to masculinize female-to-male transexuals likely produces significant metabolic dysfunction, increasing the risk factors for the development of HT and CVD, especially MetS and T2DM.
Physiological relevance and conclusions
It is increasingly apparent that endogenous TES and other androgens exert widespread beneficial effects on cardiovascular and metabolic functions. Recent human clinical trials over the last 10 years increasingly challenge the long-standing dogma that TES exerts detrimental effects on male cardiovascular and metabolic health and is largely responsible for the greater incidence of CVD in men than in women. Instead, it is now apparent that it is the gradual decline in circulating TES levels that are a normal part of the aging process that contributes to age-dependent increases in CVD and metabolic dysfunction in men. In parallel, recent experimental animal studies reveal that castration of male rats results in long-term development of HT that is completely reversed by physiological TES replacement therapy (TRT). Further, clinical hypogonadism in aging men is associated with both HT and MetS, which exacerbate the development of CVD. While the most recent human clinical trials overwhelming report that TRT does not increase risk of CVD or mortality in older hypogonadal men and is associated with reductions in BP and MetS, these studies do not provide unequivocal evidence that TRT is safe and does not increase risk of cardiovascular events. However, it is also clear that some human studies suffer from poor experimental design and statistical analysis and investigator bias. Thus, unequivocal proof that TRT is safe and beneficial for the treatment of hypogonadism and associated cardiovascular and metabolic dysfunctions in men will require more better designed clinical trials. Similarly, our understanding of the role of androgens in female health and disease is still relatively limited and more studies are needed to elucidate the mechanisms underlying the striking sexual dimorphism in the cardiovascular and metabolic effects of TES.
Acknowledgment of funding
J.N.S. acknowledges research support from the State of Texas.







Comments (0)