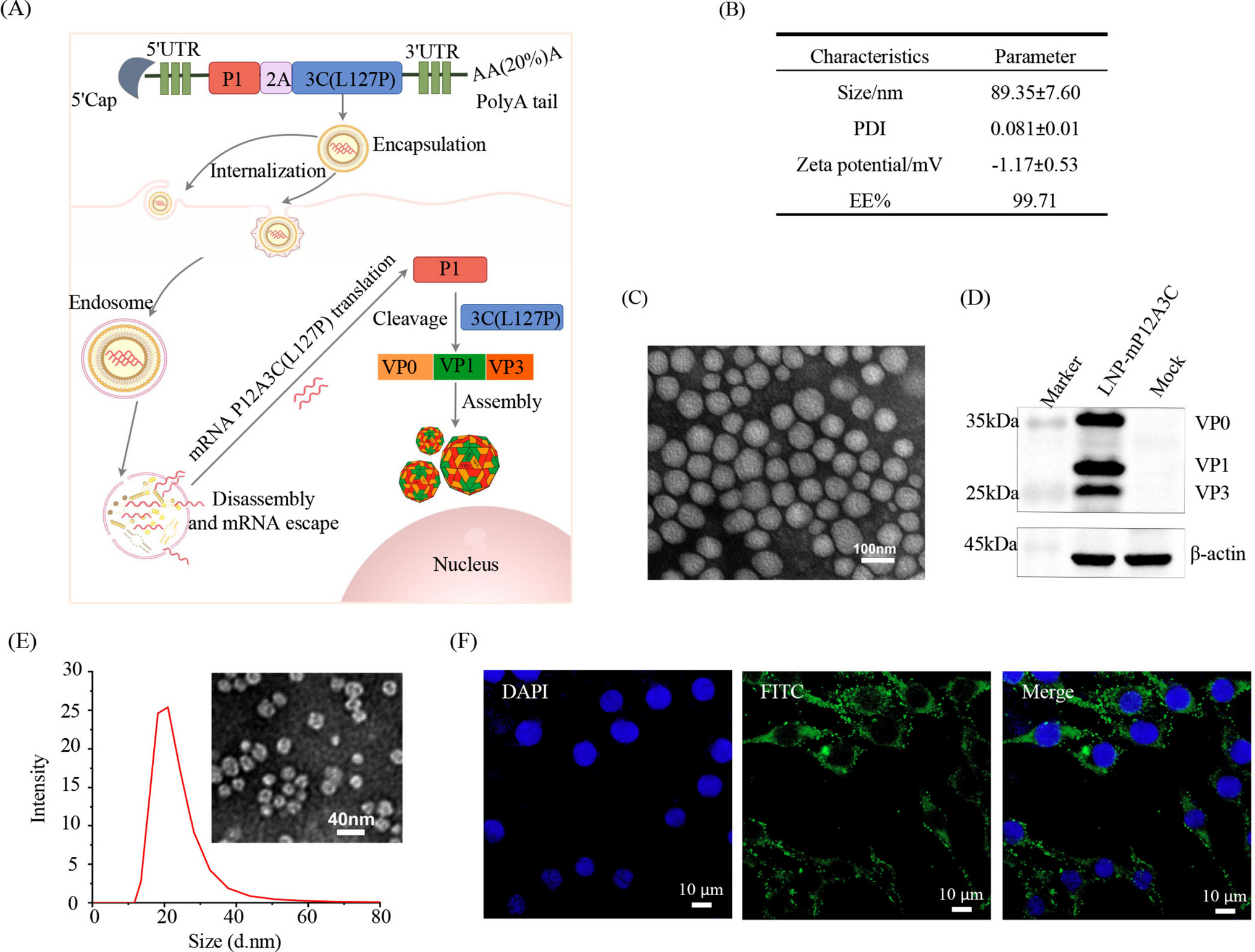
The present work describes the impact of changes in temperature and agitation on the formation of phenotypes (spore, biofilm-forming, flagellated, and lipopeptide-producing cells) of the native Bacillus paralicheniformis TB197 strain in controlled submerged cultures. Furthermore, a phenotype/genotype analysis aims to determine the correlation between these phenotypes and the production of LPs regarding both quantity and composition. The abundance of lipopeptide-producing phenotypes was estimated by analyzing the biosynthetic genes fenB and lichA, which encode the lipopeptide synthetases fengycin B and lichenysin A, respectively, since strain TB197 produces both families of LPs (Chavarria-Quicaño et al. 2023a). The expression of the tasA gene, which encodes a crucial amyloid protein for maintaining biofilm stability, was also assessed for biofilm-forming phenotypes (Cámara-Almirón et al. 2020). The hag gene, which encodes the flagellin protein, was employed to identify flagellated phenotypes, as this protein constitutes the primary structural subunit of the flagellum in Bacillus species (Molière et al. 2016). Finally, the expression of the sspB gene, which is involved in sporulation, was evaluated as a marker of this process. This gene encodes a small, acid-soluble spore protein that plays a crucial role in spore formation and stability in Bacillus species (Schoenborn et al. 2021).
The lowest temperature and agitation conditions (30 °C and 300 rpm), along with culture parameters such as aeration (1 vvm) and pH (6.5), were selected based on the most documented average conditions in the literature for lipopeptide (LPs) production by Bacillus species (Chen et al. 2015; Yeh et al. 2005; Biniarz et al. 2020; Guez et al. 2021). Considering that the B. paralicheniformis TB197 strain was isolated from a desertic area characterized by extreme environmental conditions (Chavarria-Quicaño et al. 2023b), higher temperatures (40 and 45 °C) and agitation rates (600 rpm) were also tested.
As Fig. (1) illustrates, at 30 °C, increasing the agitation rate from 300 to 600 rpm led to a 1.5-fold increase in biomass and a 2.08-fold increase in the production of LPs. It has previously been reported that high agitation in submerged culture systems promotes biomass formation and LPs production in Bacillus species, as it facilitates oxygen transfer and uniform nutrient distribution (Rangarajan et al. 2015).
However, as shown in Supplementary Fig. 6, both cases demonstrated a sharp decrease in DO (%) during the initial 12–16 h, a period characterized by bacterial exponential growth. Interestingly, the minimal DO reached at that point correlates with the time at the inflexion point (tif) of the exponential growth (Longhi et al. 2017). Afterwards, the growth rate slows down and DO begins to increase. In any case, the minimal DO remained above 30%, indicating that oxygen limitation did not occur during fermentation, as the critical oxygen threshold reported for Bacillus species ranges from 5 to 10% (Liu 2017). Remarkably, hag gene expression occurred only at 30 °C; however, at 300 rpm, it was 12.6-fold higher than that observed at 600 rpm at the same time (12 h). Although reduced, its expression continued at 24 and 48 h, while it did not persist at 600 rpm. It seems that differences in biomass production are closely related to the phenotype, with flagellated cells possibly reproducing more slowly than non-motile cells. This phenomenon, as well as LP production, appear to be influenced by shear stress rather than oxygen availability. The fenB gene and the lichA gene also responded to stressful factors, showing significant increases in expression of up to fourfold (48 h) and tenfold (24 h), respectively, higher in cultures at 600 rpm than those at 300 rpm. Additionally, the results of this study demonstrate that shear stress significantly affects the synthesis of lipopeptide variants by the TB197 strain. At 600 rpm, in addition to an overall increase in LPs production, there was a rise in LPs variant formation, with 9 distinct lipopeptides detected. In contrast, at 300 rpm, only 6 variants were identified.
Agitation and the resulting shear stress significantly affected spore formation as well (Fig. 2). At 72 h and 600 rpm, specific spore productivity (SPOSP) was nearly tenfold higher than at 300 rpm. Correspondingly, the expression of the sspB gene at 600 rpm was substantially greater at any time compared to 300 rpm (Fig. 3d). The literature on this subject is diverse and extensive. While some studies report that an increase in agitation induces sporulation (Posada-Uribe et al. 2015), others suggest that shear stress inhibits spore formation (Elisashvili et al. 2019; Lemos et al. 2015). These discrepancies may relate to the intrinsic adaptive capacity of species and even specific Bacillus strains.
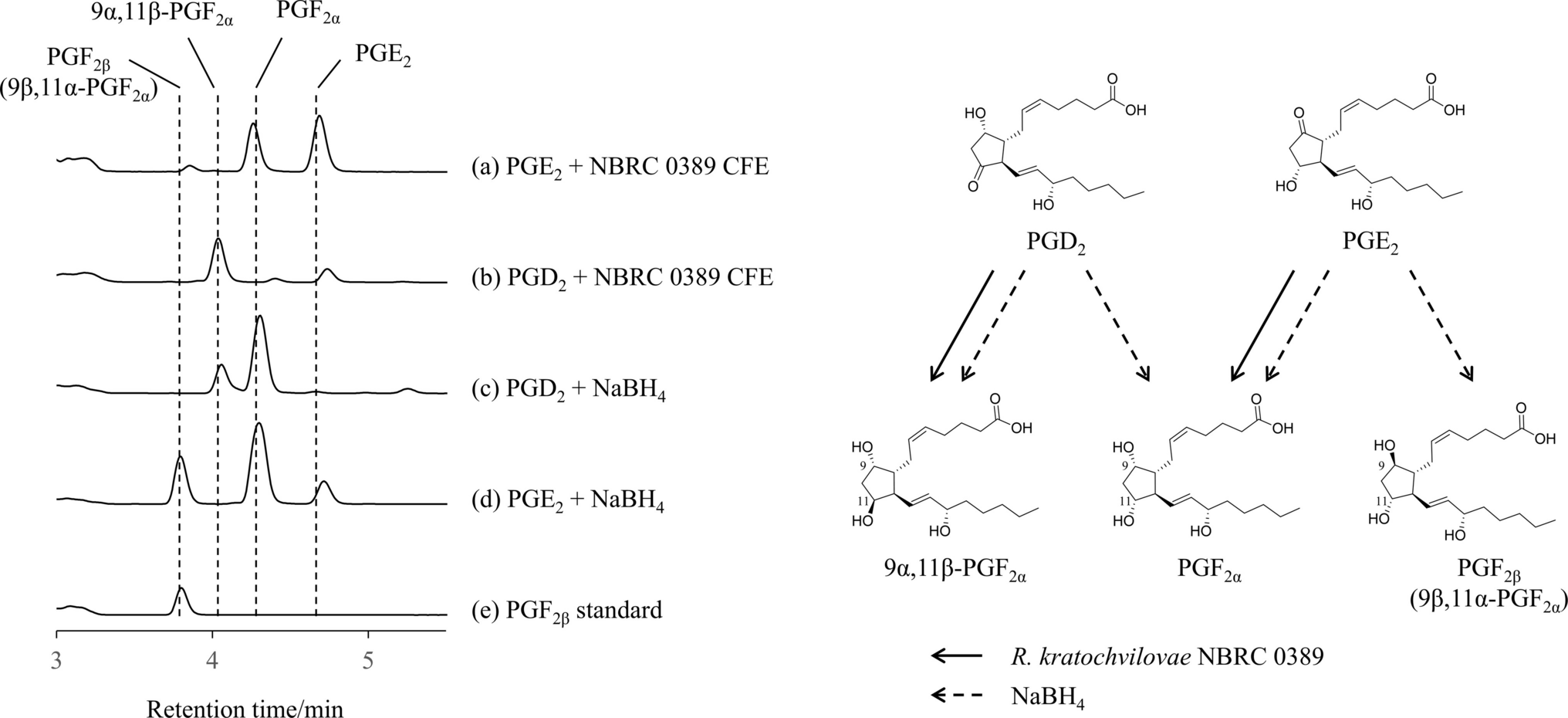
Regarding the temperature effect, the study's results demonstrated a significant increase in temperature-induced LPs production at 40 °C compared to 30 °C, with the same agitation conditions (600 rpm). Specifically, there was a 1.6-fold increase in cell mass concentration and a 2.1-fold increase in LPs titer. Notably, the LPs titers achieved at 40 °C (3.15 g/L) were remarkably high for wild-type strains (Ali et al. 2022). These findings are consistent with earlier reports (Ghribi and Ellouze-Chaabouni 2011; Coronel-León et al. 2015), which documented a correlation between increased cell growth and LPs production, suggesting that their synthesis is associated with microbial growth. Indeed, at 40 °C, the coefficient alpha was 257-fold higher than the coefficient beta (Table 2), indicating that LPs production was primarily linked to biomass growth. Interestingly, although there was no significant difference in biomass concentrations at 40 °C and 45 °C, LPs production was negatively affected at the higher temperature (2.5-fold lower), despite the α/β ratio being similar to that estimated at 40 °C (Table 2). Similarly, the estimated LPSP at 40 °C and 48 h (0.21) was threefold higher than at 45 °C at the same time (0.07). This indicates that at 45 °C, while total biomass production was similar, fewer LP-producer cell phenotypes were formed. Additionally, a clear correlation was observed between the increase in temperature and spore formation (Fig. 2), with a 19- and 42.4-fold increase in specific spore productivity (SPOSP) in cultures at 40 and 45 °C, respectively, compared to 30 °C (600 rpm).
Although the fenB gene and the lichA gene, both involved in lipopeptide biosynthesis, respond to shear stress, as previously mentioned, temperature had an even more pronounced effect on their expression, with maximum levels observed at 40 and 45 °C. Interestingly, these genes showed significant differences in their expression patterns in response to temperature and culture time (Fig. 3a and b). The highest expression of the fenB gene occurred at 12 h in cells growing at 40 °C, reaching levels 117-fold and 49-fold higher than those observed at 30 °C and 45 °C, respectively. Similarly, the highest expression of the lichA gene was found in cells cultured at 40 °C (showing no significant differences based on culture time), approximately 18-fold greater than in cells growing at 45 °C (with no significant differences according to culture time) and 38-fold higher than those at 30 °C (600 rpm) after 12 or 24 h of culture. In contrast to cultures at 40 and 45 °C, where the expression of the lichA gene remained unchanged throughout fermentation, at 30 °C, the relative expression level dropped eightfold at 48 and 72 h.
The factors regulating the expression of fengycin synthase genes are an underexplored area of research. It is known that the pleiotropic regulator DegQ induces the expression of genes related to fengycin synthesis, and that its overexpression correlates with increased production of this lipopeptide (Wang et al. 2015). The DegQ protein is part of the quorum sensing system in Bacillus, and its expression is induced in response to environmental stress (Wang et al. 2015). This suggests that stressful factors such as high temperatures and shear, associated with high agitation rates (40 °C and 600 rpm), induce DegQ expression in LP-producer cells, leading to enhanced fengycin production. This phenomenon was previously observed by Ha et al. (2018), who reported a 1.36-fold increase in surfactin production when agitation was increased from 300 to 500 rpm. However, at 45 °C, other factors, such as spores and biofilm-forming cells, appear to govern the system.
Lichenysin synthases are responsible for synthesizing lichenysins and surfactins (Tapi et al. 2010), which are secondary metabolites that, unlike primary metabolites, are not essential for cell survival. However, the high and consistent expression of the lichA gene at elevated temperatures suggests that these compounds may be crucial for cell survival under stress conditions. While further validation is needed to support this hypothesis, surfactins, in addition to their inhibitory and competitive functions, have been shown to act as signaling molecules in quorum sensing systems, promoting cell differentiation under stress (Qi et al. 2023; Schoenborn et al. 2021). The results of this study indicate that molecules belonging to the lichenysin family may also act as cell signaling molecules, as a higher abundance of biofilm- and spore-forming phenotypes was observed in cultures at 45 °C (Figs. 2 and Supplementary Fig. 5).
Interestingly, the results of this study show that temperature modulates the synthesis of lipopeptide variants by the TB197 strain as well. At 40 °C, alongside an overall increase in LPs production, a rise in LPs variant formation was observed, with 14 distinct lipopeptides detected. Conversely, at 30 and 45 °C (600 rpm), only 9 and 13 variants were identified, respectively (Table 3, Supplementary Fig. 2). Despite the relatively limited number of variants identified in these treatments, some interesting observations should be noted. For example, a new potential variant of the fengycin family (peak 3; Table 3 and Supplementary Fig. 2), namely fengycin A C19, was evident (Mnif et al. 2023). Our previous studies did not detect this variant (Chavarria-Quicaño et al. 2023a). Additionally, the C12 variant of the lichenysin family was unique to the 40 °C treatment. It has been previously reported that specific carbon sources may influence the production of certain LPs variants over others (Bartal et al. 2018). However, there is currently no evidence to suggest that agitation and temperature play a role in the synthesis or inhibition of specific LPs variants. Therefore, this study contributes novel insights to the field, as it is the first to demonstrate that temperature can affect the synthesis and diversification of lipopeptide variants in controlled liquid cultures of strain TB197.
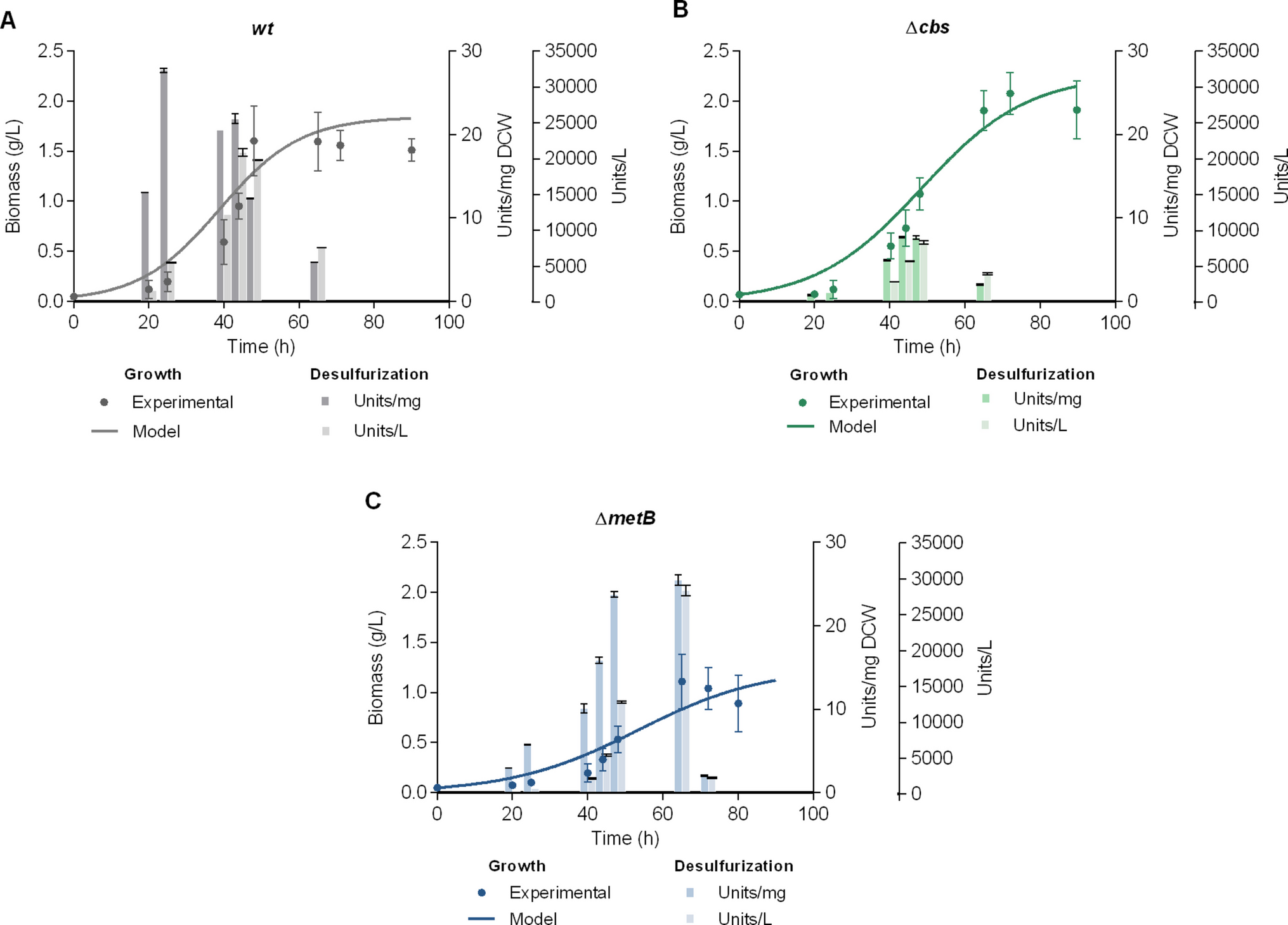
On the other hand, to evaluate the prevalence of biofilm-producing phenotypes, the relative expression of the tasA gene was analyzed (Fig. 3c). The expression of the tasA gene was strongly induced by temperature, showing a 4.3- and 33-fold increase at 40 and 45 °C, respectively, compared to 30 °C (600 rpm; 12 h). These genotypic results suggested that higher temperatures, such as 45 °C, promoted biofilm formation (a phenotypic response), visible on the bioreactor walls and agitation components as abundant, yellowish, robust structures (Supplementary Fig. 5). Other researchers have previously documented the impact of temperature on biofilm formation in Bacillus species (Liu et al. 2023). However, most studies have primarily focused on biofilm formation in solid cultures, emphasizing the characteristics of colonies and biofilms. Elhariry (2014) reported that B. coagulans and B. stearothermophilus strains exhibited enhanced biofilm formation at 45 °C when grown on solid media. The resulting colonies were significantly larger and more robust than those formed at 37 °C. A comparable phenomenon was documented by Rajitha et al. (2021) in B. haynesii strains cultivated at 45 °C, where enhanced amyloid protein formation and increased colony size and roughness were observed.
In alignment with these findings, the results of this study illustrate that elevated temperatures in submerged cultures in controlled bioreactors also stimulate biofilm formation. It is important to note that even at 30 °C and 40 °C, the expression of the tasA gene was observed; however, the phenotypic response was minimal, with limited biofilm formation, unlike the results at 45 °C (Supplementary Fig. 5). This outcome may be attributed to the fact that, although tasA was used as a marker gene associated with biofilm-forming phenotypes, it is not the only gene involved in this complex process. Biofilm formation is a multifactorial phenomenon involving multiple genes and regulatory networks; therefore, tasA expression alone does not necessarily ensure the structural development of a biofilm (Diehl et al. 2018). These findings highlight the need for further studies in agitated liquid cultures to better understand the dynamics of gene expression and their relationship with biofilm morphogenesis under varying environmental conditions.
As the tasA gene, the relative expression of the sspB gene, which is related to spore-forming cells, displayed a strong response to stressful factors such as high temperatures (40 and 45 °C; 600 rpm) (Fig. 3d), particularly between the later exponential (24 h) and earlier stationary (48 h) phases. The most significant response occurred at 45 °C and 48 h, being 3.6- and 15.8-fold higher than the maximum relative expression of the sspB gene observed at 24 h in cultures at 40 °C and 30 °C (600 rpm), respectively. As indicated by previous genotypic results, the relative expression patterns of the sspB gene strongly support the phenotypic response shown in Fig. 2. Additionally, Isticato et al. (2020) showed that elevated temperatures enhance sporulation and alter spore structure and functionality, reinforcing our findings. Sporulation in Bacillus species serves as a resistance mechanism typically activated in response to nutrient starvation and abiotic stress, often manifesting during the stationary phase (Elhariry 2014). However, our results indicate that the relative expression of the sspB gene begins as early as the mid-exponential phase (12 h). Since sporulation is an energetically expensive process and is generally considered the final survival strategy, this premature activation suggests that the conditions evaluated were, in some instances, stressful, prompting the cells to initiate sporulation at early growth stages.
Finally, as previously mentioned, relative gene expressions linked to flagellated phenotypes (hag) were only detected in cultures grown at 30 °C (Fig. 3e). The lack of hag gene expression at the highest temperatures (40 and 45 °C) shows that heat stress completely suppresses its expression and, consequently, the development of flagellated phenotypes.
The formation of flagella and biofilms represents a significant energetic cost for Bacillus cells; therefore, a high level of regulation governs the development of these phenotypes (Bucher et al. 2015). Steinberg et al. (2020) indicate that a regulatory switch in Bacillus cells allows for the normalization of biofilm production and repression of motility and vice versa. This regulatory switch primarily depends on two master regulators: the homologous proteins SinR and SlrR. During Bacillus’s planktonic growth, SinR represses the expression of biofilm-forming operons (epsA-O and tapA-sipW-tasA) and the expression of SlrR, leading to the promotion of flagellated phenotypes (Vlamakis et al. 2008). Conversely, once biofilm formation is initiated, the SlrR protein inhibits SinR activity by binding to it and preventing SinR from binding to DNA, thus activating the operons associated with biofilm formation. SlrR, in turn, binds to SinR, forming a heterodimer that represses the fla/che operon, which encodes essential motility components (Steinberg et al. 2020). Other explanations may be associated with the high LPs titers observed at 40 °C, where hag gene expression was undetectable. It has been previously demonstrated that LPs belonging to the surfactin family can inhibit the expression of genes involved in motility (Bin et al. 2021). Many Bacillus species secrete surfactin, reducing the surface tension between bacterial cells and substrates, thereby facilitating bacterial movement on surfaces without flagella (Bin et al. 2021; Luo et al. 2015).
The main results of this work are summarized as follows: (1) Increased agitation and temperature stimulate the growth of the TB197 strain and, to some extent, the production of LPs. (2) The kinetic analysis conducted at 40 and 45 °C revealed comparable biomass concentrations; however, notable differences were observed in LPs production, linked to variations in the cell-phenotype composition of the biomass. At 40 °C, the LPs titer was 3.14 g/L, suggesting that this temperature may promote a higher abundance of LP-specialized phenotypes. (3) Exposure to elevated temperatures induces the formation of sporulating and biofilm-forming phenotypes in strain TB197, indicating adaptive mechanisms that enable this strain to thrive under such conditions. Interestingly, the relative expression analyses of genes associated with specific phenotypes correlated closely with the biomass and LPs production kinetic results, as well as LPs composition. However, since the increase in relative expression level did not correlate with a linear physiological response (e.g., relative expression level of LP-related genes vs LPs titers), translational and/or post-translational regulatory systems and/or even negative feedback loops in enzyme activity may also be involved.


Comments (0)