
Remember me

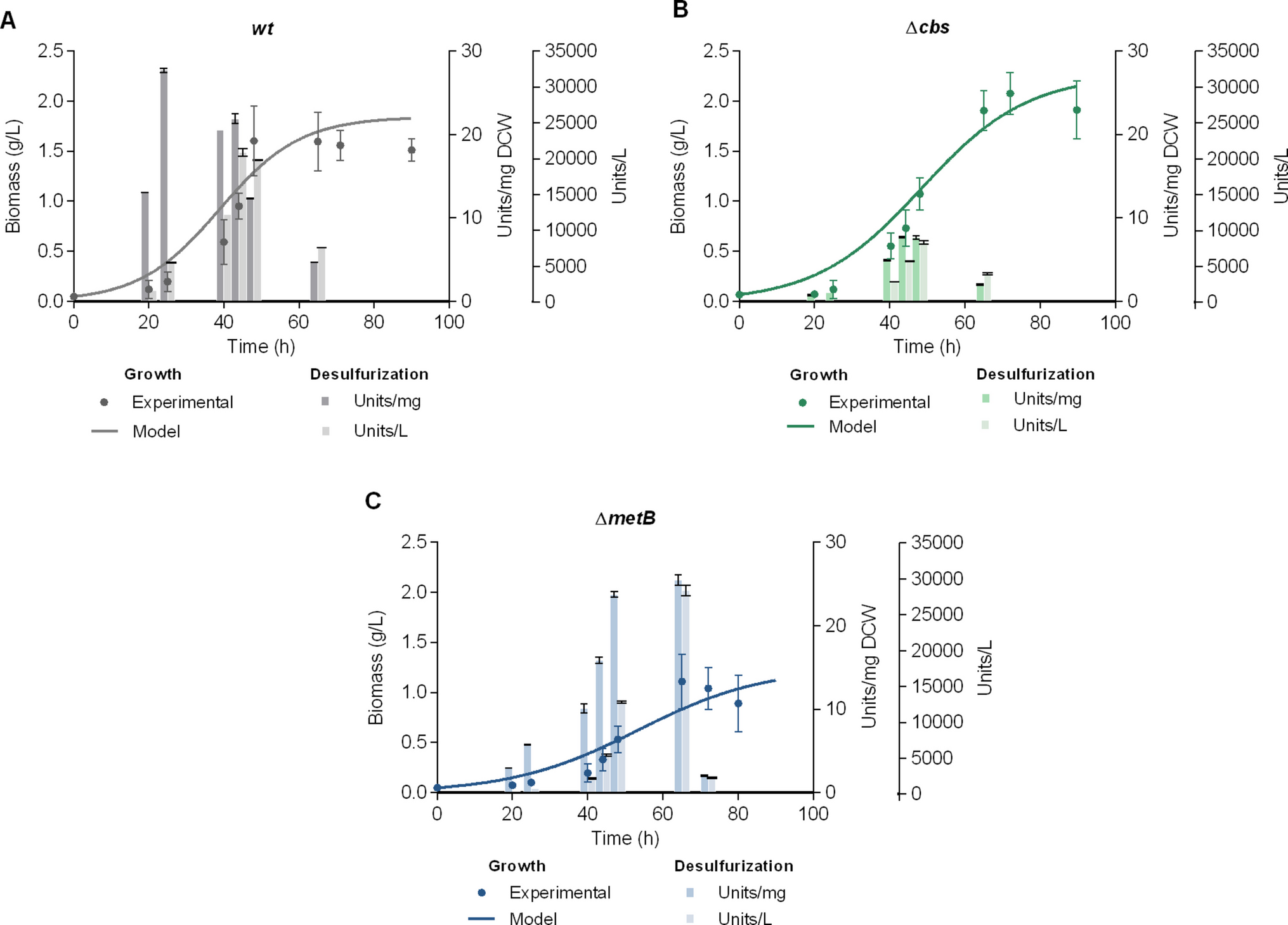
To investigate the desulfurization potential of Δcbs and ΔmetB recombinant R. qingshengii IGTS8 strains at the bioreactor scale, an initial assessment of growth and resting-cell desulfurization activity was performed in 5 L batch cultures under non-repressive conditions, with the wild-type (wt) strain included for comparison. Each strain was cultivated in the presence of its previously determined optimal sulfur source (1 mM). Dimethyl sulfoxide (DMSO) was used for the wt strain, as methionine, cysteine, and under certain medium compositions, sulfate, are known to repress its desulfurization activity. Sulfate was used for the Δcbs strain, and methionine for the ΔmetB strain (Fig. 1) (Li et al. 1996; Martzoukou et al. 2022, 2023b). Ethanol (165 mM) was selected as the carbon source for all strains, based on its proven efficacy in promoting growth and desulfurization activity in both wt and recombinant R. qingshengii IGTS8, even in the presence of different culture media (Aggarwal et al. 2011; Martzoukou et al., 2022, 2023a, b, 2025). The maximum biomass yield for the wt strain was equal to 1.60 ± 0.35 gDCW/L at 48 h of growth, and the maximum specific desulfurization activity was documented at 25 h of growth (27.68 ± 0.24 Units/mgDCW, Fig. 1A), albeit in small-scale cultures the respective BDS maximum was documented at the mid-log phase (45 h; 29.03 ± 1.13 Units/mgDCW) (Martzoukou et al. 2022). In addition, a reduction of desulfurization activity was observed across the stationary phase. Concurrently, the ethanol consumption curve demonstrated that the carbon source is completely depleted after 48 h, i.e., at the end of the exponential phase in batch cultures, which may account for the reduction of desulfurization activity and the initiation of stationary/death phase (Supplementary Information, Fig. S2). The maximum volumetric activity for the wt strain was documented at the exponential phase, prior to the depletion of ethanol (44 h; 20848 ± 556 Units/L) (Fig. 1A). The recombinant strain Δcbs exhibited the maximum biomass yield (2.08 ± 0.21 gDCW/L) at 72 h of growth, the maximum specific desulfurization activity at 44 h (7.71 ± 0.07 Units/mgDCW) and the maximum volumetric desulfurization activity at 48 h (8250 ± 251 Units/L) (Fig. 1B). In small-volume cultures of the same strain and under the same medium composition, the specific BDSmax was also documented for cells harvested in the mid-log phase and found equal to 15.23 ± 0.27 Units/mgDCW (Martzoukou et al. 2022). Comparison of the two experimental setups in the case of Δcbs underscores a significantly enhanced biomass yield for the bioreactor-scale culture (P < 0.01; Biomass microplate: 1.09 ± 0.03 gDCW/L versus Biomass bioreactor: 2.08 ± 0.21 gDCW/L), which however resulted in a significant decrease of specific desulfurization activity compared to the 96-well plate culture (P < 0.001). The ΔmetB recombinant strain exhibited maximum biomass yield (1.11 ± 0.27 gDCW/L) and maximum specific and volumetric desulfurization activities (25.46 ± 0.65 Units/mgDCW and 28261 ± 716 Units/L, respectively) for cells harvested at 65 h of growth (Fig. 1C), in contrast to the wt and Δcbs strains that reached maximum catalytic activity at earlier growth stages. However, within-strain comparison of the specific desulfurization activities (Units/mgDCW) at 48 and 65 h for ΔmetB indicated only marginal differences, whereas the volumetric desulfurization activity (BDSvol, in Units/L; Fig. 1C) appears significantly enhanced at the end of the exponential phase, a fact attributed to the observed increase in biomass concentration (P < 0.0001; ΔmetB; BDSvol 48 h versus 65 h). Small-volume microplate cultures of ΔmetB, in the presence of 1 mM methionine in the same medium, yielded a BDSmax equal to 15.40 ± 0.13 Units/mgDCW (Martzoukou et al. 2022). Batch cultures of ΔmetB at the bioreactor-scale exhibited significantly enhanced maximum desulfurization activity compared to previously-conducted microplate cultures (P < 0.001) while both maxima were documented at the late-log phase (65 h) (Martzoukou et al. 2022). Notably, comparison between the two different growth setups for each strain revealed an enhancement in specific BDS activity only for the ΔmetB recombinant strain, whereas the maximum desulfurization activity remained unaffected for the wt strain (P > 0.05), and was significantly reduced for the Δcbs recombinant (P < 0.001), indicating a strain-specific response to the increased culture volume and to the transition from a 96-well plate to a 5 L bioreactor with controlled aeration and pH (Martzoukou et al. 2022 compared to the present study). The recombinant strain ΔmetB, when grown in the presence of methionine (normally repressive for wt) exhibited maximum specific desulfurization activity comparable to that of the wt strain grown in the presence of the non-repressive sulfur source DMSO (1 mM sulfur; P > 0.05; ΔmetB, 65 h, 25.46 ± 0.65 Units/mgDCWversus wt, 25 h, 27.68 ± 0.24 Units/mgDCW). The sulfate-grown Δcbs recombinant strain exhibited the lowest desulfurization activity, reaching a maximum of only 7.71 ± 0.07 Units/mgDCW at 44 h (P < 0.0001; BDSmax; Δcbs versus wt, ΔmetB). Comparison of volumetric desulfurization activities highlighted a statistically significant maximum for the ΔmetB strain at 65 h (1 mM sulfur; P < 0.001 for ΔmetB, 65 h, 28261 ± 716 Units/L versus wt, 44 h, 20848 ± 556 Units/L).
Fig. 1 The alternative text for this image may have been generated using AI.
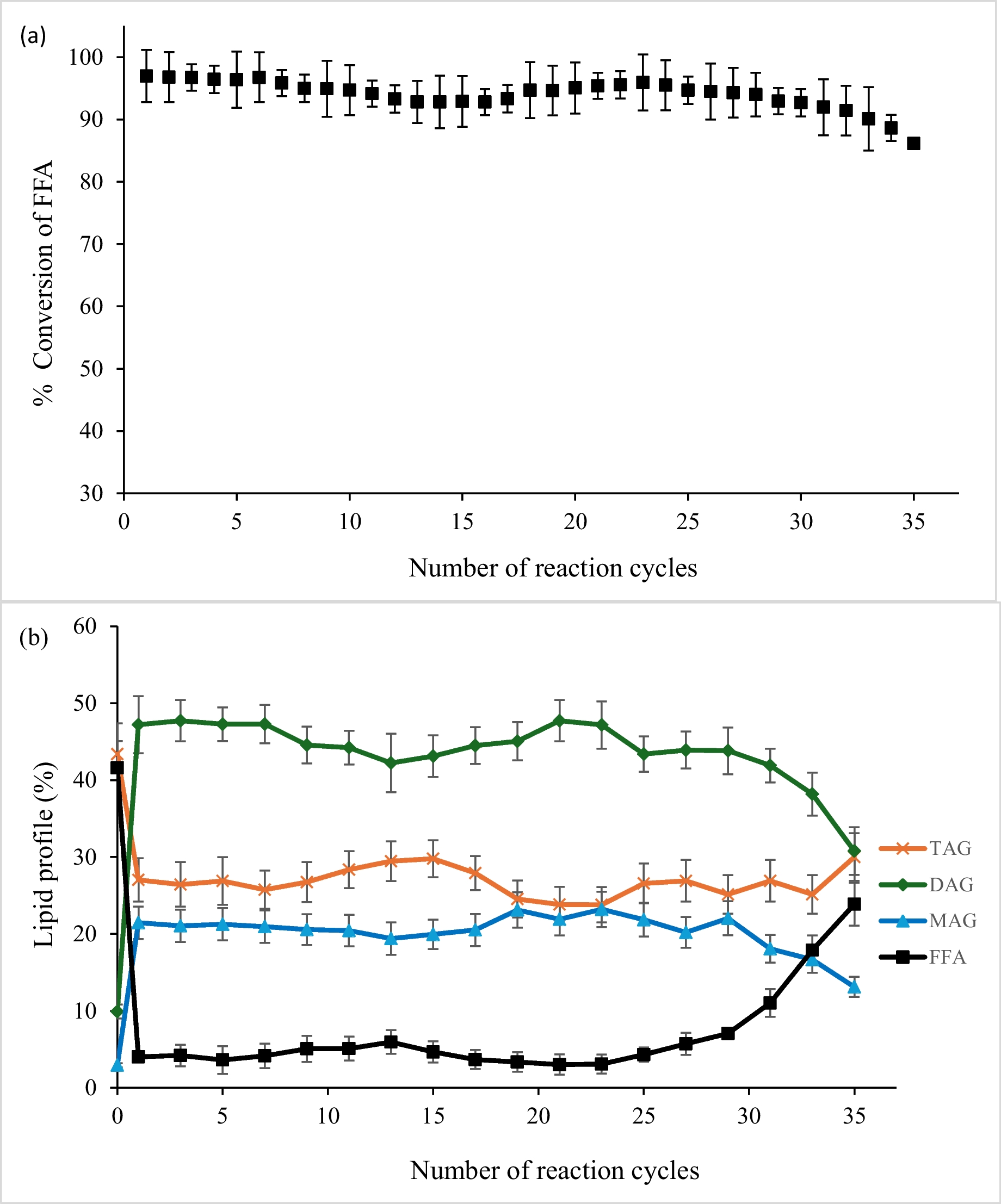
The alternative text for this image may have been generated using AI.Comparison of the biodesulfurization activities of wt and recombinant strains grown in the presence of their optimum sulfur sources. Growth (Biomass, g/L), specific desulfurization activity (Units 2-HBP/mg Dry Cell Weight [DCW]) and volumetric desulfurization activity (Units 2-HBP/L) of wt A, Δcbs Β, and ΔmetB C strains. All cultures were supplemented with 165 mM ethanol as the sole carbon source. DMSO, sulfate, or methionine (1 mM sulfur) were used as the sole sulfur source for strains wt, Δcbs, and ΔmetB, respectively. The volume of batch cultures was 5 L in all cases
The growth kinetic parameters Cmax (maximum calculated biomass, gDCW/L), and μmax (apparent maximum specific growth rate, h−1 (g/L)−1) were obtained by fitting the biomass concentration versus time experimental values to the logistic equation for all strains and are presented in Table 2.
Table 2 Growth kinetic parameters calculated for wt and recombinant R. qingshengii IGTS8 strains. All cultures were supplemented with 165 mM ethanol as the sole carbon source. DMSO, sulfate, or methionine (1 mM sulfur) were used as the sole sulfur source for strains wt, Δcbs, and ΔmetB respectively. Kinetic parameters were calculated by fitting experimental values for biomass concentration (g/L) versus time (h), to the logistic model. See also Fig. 1. Values represent mean ± SEMNotably, Δcbs exhibited the highest Cmax among the three strains, although the results were non-significant, whereas the biomass yield of Δcbs was significantly higher than that of ΔmetB (P < 0.01). Contrastingly, ΔmetB exhibited marginally lower values for the calculated growth kinetic parameters compared to the wt strain (P > 0.05), and a significantly lower biomass yield (P < 0.05), an effect which has been observed previously for small-scale cultures of both recombinant strains (Δcbs, ΔmetB) in the presence of methionine (Martzoukou et al. 2022).
The comparative analysis of bioreactor performance revealed that while Δcbs achieved higher biomass yields, the superior enzymatic activity of ΔmetB resulted in greater overall desulfurization capacity per culture volume. Notably, ΔmetB maintained desulfurization activity levels comparable to the wt strain despite utilizing methionine, an otherwise repressive sulfur source. Medium optimization of large-scale cultures can result in enhanced desulfurization rates, higher biomass yield, and/or prolonged desulfurization activities, and thus the effect of varying methionine and ethanol concentrations was further explored for the ΔmetB strain
Effect of methionine concentration on ΔmetB batch culturesIn the context of bioreactor-scale culture medium optimization studies, we initially investigated the effect of sulfur source concentration on ΔmetB strain growth and desulfurization activity. Methionine was supplemented at 1, 2, and 4 mM as the sole sulfur source in 5 L bioreactor batch cultures, while 165 mM ethanol was utilized as the sole carbon source in all cases (Fig. 2). Statistical comparison of the obtained biomass yield (1.06–1.12 gDCW/L), maximum calculated biomass (Cmax range: 1.10–1.25 gDCW/L, Table 3) and maximum specific desulfurization activity (\(}_}^}\) range: 25.00–25.47 Units/mgDCW, Table S1) did not reveal any significant differences in the presence of the three sulfur concentrations tested (P > 0.05). However, volumetric biodesulfurization activity was significantly higher in the presence of 1 mM methionine, compared to the other two supplemented sulfur concentrations (P < 0.001). Notably, 2 mM methionine resulted in the highest growth rate (μmax = 0.08 h−1 (g/L)−1) compared to 1 and 4 mM supplemental methionine (Table 3), and thus, the maximum biomass yield is achieved faster, while the specific desulfurization activity is maintained at high levels throughout the logarithmic growth phase of the culture.
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Effect of methionine concentration on ΔmetB strain growth and resting-cell desulfurization activity. Growth (Biomass, g/L), specific desulfurization activity (Units 2-HBP/mg Dry Cell Weight [DCW]) and volumetric desulfurization activity (Units 2-HBP/L) of ΔmetB in the presence of 1 mM A, 2 mM Β, and 4 mM C methionine as the sole sulfur source. All cultures were supplemented with 165 mM ethanol as the sole carbon source. The volume of batch cultures was 5 L in all cases. Please note that Fig. 1C is repeated in A
Table 3 Kinetic parameters calculated for ΔmetB strain grown in different culture conditions. Ethanol and methionine were supplemented as the sole carbon and sulfur sources, respectively. See also Fig. 2, Fig. 3Effect of ethanol concentration on ΔmetB batch culturesCarbon source concentration is known to exert an effect on R. qingshengii IGTS8 growth kinetic parameters, while time-dependent variations in desulfurization efficiency have also been reported (Martzoukou et al. 2023a). The effect of ethanol concentration (45, 87, and 165 mM) on ΔmetB recombinant strain performance was examined using 2 mM methionine as the optimal sulfur source (Fig. 3). Maximum specific desulfurization activity (\(}_}^} )\) was not significantly affected by the carbon content (Table S1), albeit at 45 mM ethanol a slightly reduced average desulfurization efficiency was observed (P > 0.05) (Fig. 3 and Table 3). Maximum biomass yield significantly decreased with lower ethanol concentrations (P < 0.01, for 45 mM versus 87 mM EtOH; P < 0.01, for 87 mM versus 165 mM EtOH; P < 0.0001, for 45 mM versus 165 mM EtOH). Furthermore, a correlation was observed between the carbon content and culture duration. For instance, at an ethanol concentration of 165 mM and 2 mM methionine, the maximum biomass yield was attained after 49 h of growth, compared to 40 h for lower concentrations. Volumetric activity (Units/L) was also significantly reduced at 45 mM ethanol (P < 0.0001 versus 87 and 165 mM EtOH).
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Effect of ethanol concentration on ΔmetB strain growth and resting-cell desulfurization activity. Growth (Biomass, g/L), specific desulfurization activity (Units 2-HBP/mg Dry Cell Weight [DCW]) and volumetric desulfurization activity (Units 2-HBP/L) of ΔmetB in the presence of 45 mM A, 87 mM Β, and 165 mM C ethanol as the sole carbon source. All cultures were supplemented with 2 mM methionine as the sole sulfur source. The volume of batch cultures was 5 L in all cases. Please note that Fig. 2B is repeated in C
The effect of ethanol concentration was also investigated in the presence of 1 mM methionine. Under these conditions, biomass yield was reduced 2.6-fold, which can be attributed to the lower carbon availability in the medium caused by the ethanol concentration used (P < 0.001, 45 mM versus 165 mM EtOH) (Fig. S3 and Table 3). Of note, the culture supplemented with the lower ethanol and methionine concentrations, was also characterized by reduced logarithmic phase duration (40 h, 45 mM EtOH and 1 mM methionine). Table 3 and Table S1 summarize the calculated kinetic parameters and the maximum biodesulfurization activities, respectively, for the ΔmetB strain grown in the presence of different ethanol and methionine concentrations. Biomass concentrations and culture duration increased with the supplementation of higher ethanol concentrations, whereas the increase in methionine concentration (2 mM) only marginally affected growth rate (μmax). Volumetric desulfurization activity exhibited a significant increase in correlation with the carbon concentration. Overall, optimal culture conditions were established at 165 mM ethanol and 2 mM methionine, which balanced biomass production with enzymatic activity, while lower carbon concentrations led to reduced biomass.
Investigating the effect of ethanol feed rateA typical problem often observed in batch cultures is the reduction of specific desulfurization activity as the cells exit the logarithmic growth phase, a phenomenon that coincides (but not necessarily correlates) with the depletion of carbon source (Fig. S2). In this context, we assessed the effect of varying ethanol (carbon source) feed rates in fed-batch cultures, in order to investigate the possibility of optimizing ΔmetB growth characteristics and improving the stability of their desulfurization activity. We used CDM culture medium containing 2 mM methionine as the sole sulfur source. Experiments were initiated as batch cultures at an initial volume V0 = 3L and initial ethanol concentration of 165 mM (7.6 g/L). Ethanol feed (in CDM medium containing 2 mM methionine) was applied after 46 ± 2 h, a time point coinciding with the logarithmic phase and a point up to which the desulfurization activity is relatively stable, following what was observed in the corresponding batch process. Feeding continued until the total volume of the culture exceeded 5.5 L. In all cases we monitored culture growth, desulfurization activity, and ethanol levels.
Initially we applied a medium feed (FF) of 19.2 mL/h at an ethanol (carbon source) concentration of 1 M (46.1 g/L) that corresponds to an ethanol feed of 0.89 g/h (Fig. 4A and D). Under this feeding regime, ethanol concentration in the culture supernatant became undetectable after 60 h (approx. 15 h post-feeding initiation). In an effort to maintain a detectable ethanol concentration in the culture, we tried a second fed-batch experiment keeping all parameters identical but increasing FF to 28.2 mL/h (1.30 g ethanol/h). Notably, this time (Fig. 4B and E) cell growth was unable to consume the added ethanol which started to increase gradually exceeding 10 g/L at the final stages of the experiment. In an effort to optimally control ethanol levels in the fed-batch culture, a third experiment was performed where we aimed for an intermediate value of ethanol feed rate. In this case, we slightly increased the FF to 30.6 mL/h while reducing ethanol feed concentration to 0.8 M (36.8 g/L), thus achieving an ethanol feed of 1.13 g/h (Fig. 4C and F). At this feeding regime, it was possible to achieve some kind of dynamic equilibrium between ethanol feed rate and consumption, that maintained the ethanol concentration in the bioreactor at levels around 5 g/L. Analysis of the three fed-batch cultures indicated a significant improvement in the biodesulfurization activity of the cells for the third feeding regime. Specific BDS activity was notably elevated during the feeding phase, which contributed to sustained high overall activity in the bioreactor throughout the duration of the experiment (P < 0.01, \(}_}^}\) F3versus F1) (Fig. 4A – C and Table S2). In F1 and F2, specific BDS reached the maximum at ~ 50 h of growth and gradually declined thereafter. Contrastingly, in F3 high BDS values were documented for up to 72 h, in addition to achieving the highest overall values for volumetric and total desulfurization activities (P < 0.01 for \(}_}^}\) and \(}_}^}\), F3versus F2) (Fig. 4D – F and Table S2). Statistical analysis of the experimentally measured biomass concentration highlights a significantly enhanced yield for the feed rate of 0.89 g/h, compared to 1.13 and 1.30 g/h (P < 0.01). The highest biomass value (in grams) was achieved for the F1 and F3 fed-batch cultures (13.5–13.6 gDCW; P < 0.0001 compared to F2). Importantly, the intermediate ethanol feed rate (F3) resulted in the highest units of desulfurization activity, whereas F1 and F3 both resulted in the highest biomass yields.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Fed-batch cultures of ΔmetB strain with varying ethanol feed rates. A–C Growth (Biomass, g/L), ethanol concentration (g/L), and resting-cell specific desulfurization activity (Units 2-HBP/mg Dry Cell Weight [DCW]) for feed rates of F1 = 0.89 (A), F2 = 1.30 (Β), and F3 = 1.13 g ethanol/h (C), listed in the order in which they were investigated. D–F Growth (Biomass, gDCW), ethanol consumption (g), and resting-cell total desulfurization activity (Units 2-HBP/L ∙ culture volume, in L) for the same feed rates as in (A-C), respectively. All culture media were supplemented with 2 mM methionine as the sole sulfur source. The volume of initial batch cultures was 3 L and the initial concentration of ethanol was 165 mM (7.6 g/L). The fed-batch processes were initiated at 45, 48, and 46 h, respectively. See also Table S2
Of more industrial relevance are the comparison data that appear in Fig. 5. Maximum volumetric BDS activities for ΔmetB strain were markedly increased in all fed-batch cultures when compared to the batch culture (Fig. 5A). For fed batches F1 and F3, values doubled from 21137 Units/L at 49 h in the batch culture to 42117 Units/L at 72 h in fed-batch F3. This corresponds to an almost 40% increase in the total BDS productivity (Units/L/h) at the time-point of maximum volumetric activity, and way higher at later time points. The cellular physiological variations among the different ethanol feeding rates are also reflected in the total desulfurization activity per grams of ethanol consumed. Feeding regime F3, where the ethanol concentration was practically maintained constant, proved superior by maintaining the highest volumetric BDS activities and yields with respect to ethanol consumption, throughout the feeding part of the cultures (Fig. 5B).
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.A Volumetric desulfurization activity (Units/L) as a function of time (h), and B Total desulfurization activity per grams of ethanol consumed, for fed-batch cultures of ΔmetB strain. The carbon source feeds were 0.89, 1.30, and 1.13 g ethanol/h, and the flow rates were 19.2, 28.2, and 30.6 mL/h for F1 – F3, respectively. Methionine (2 mM) was the sole sulfur source. The volume of initial batch cultures was 3 L and the initial concentration of ethanol was 165 mM (7.6 g/L). The fed-batch processes were initiated at 45, 48, and 46 h, respectively. The switch to fed-batch is indicated with vertical grided lines. The batch shown in (A) was supplemented with 2 mM methionine and 165 mM ethanol
Comparison of the fed-batch cultures with the batch culture of the same initial carbon and sulfur concentrations, i.e., 165 mM ethanol and 2 mM methionine, highlights a significant enhancement in catalytic activity, in terms of maximum specific, volumetric, and total desulfurization activities (\(}_}^} :\) P < 0.01, F1versus batch culture; P < 0.001, F2 and F3versus batch; Table S2). Notably, the volumetric desulfurization activity was up to 99.3% higher for the fed-batch cultures (\(}_}^}\): P < 0.001 for F1 and F3versus batch; P < 0.01 for F2versus batch; Table S2), and the total desulfurization activity was up to 51.3% higher (\(}_}^} :\) P < 0.05 for F1 and F3versus batch; Table S2).
Comments (0)