
Remember me

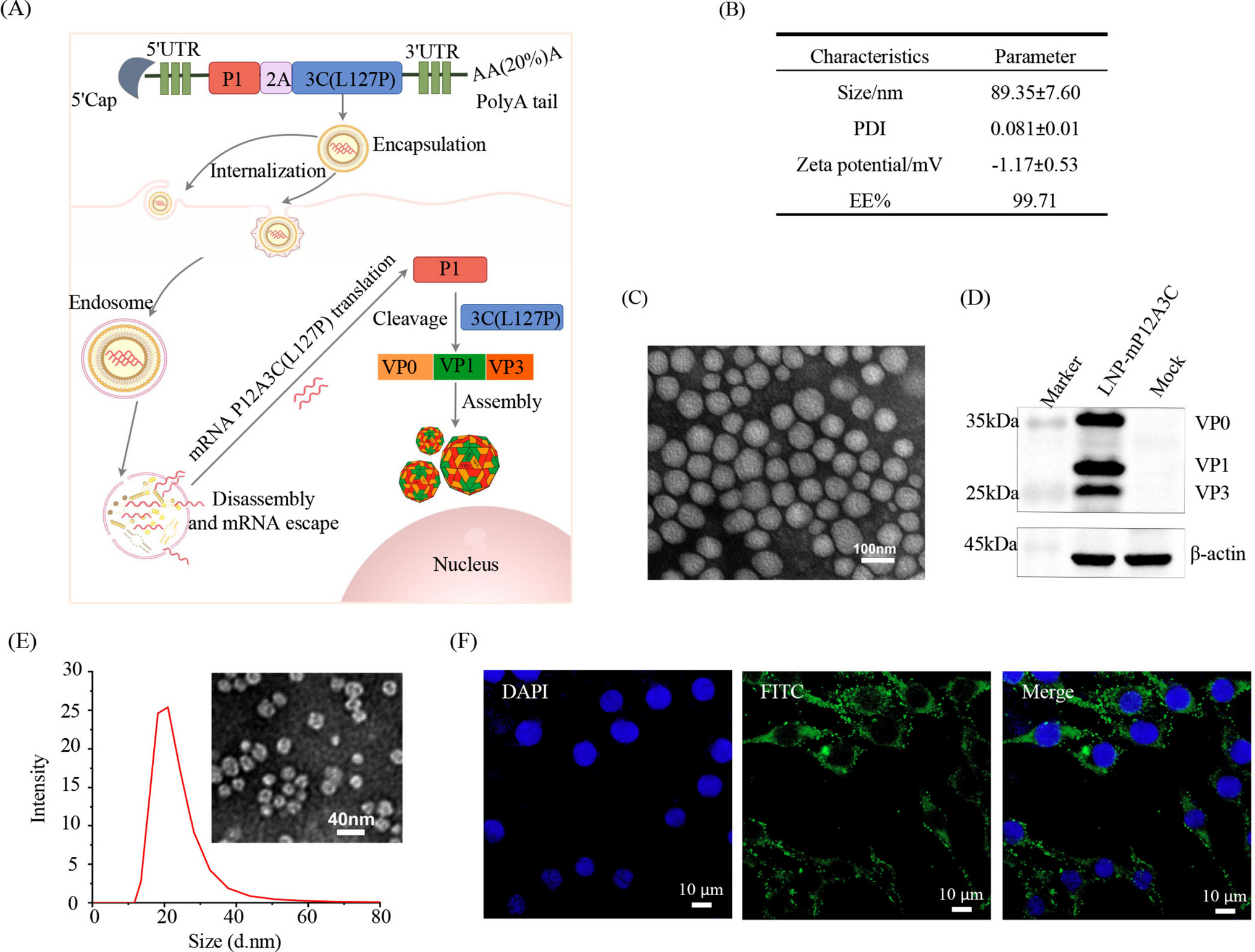
To investigate the spontaneous genomic alterations of Y. lipolytica, nine wild-type isolates derived from strain PO1f (Table 1) were subcultured on solid YPD medium for 85 generations to accumulate mutations (Fig. 1A). Whole-genome sequencing of these nine isolates identified 62 SNVs (Dataset S1-1) and 37 InDels (Dataset S1-2). Using the formula described in the “Materials and methods” section, we calculated that spontaneous rates of SNVs and InDels were 1.8 × 10−10 (62/9/25/85/20,500,000) and 9.4 × 10−11 (37/9/25/85/20,500,000) per base per cell division, respectively (Fig. 1B). These values are comparable to those previously reported for the wild-type Y. lipolytica strain with a W29 genetic background (Xiong et al. 2025), suggesting minimal strain-specific variation in spontaneous mutation rates. C:G to T:A (C to T or G to A) and C:G to A:T were the prominent types of base substitution, accounting for up to 42% and 25% of all SNVs, respectively (Fig. 1C). C:G to T:A substitutions commonly arise from the spontaneous deamination of 5-methylcytosine (5mC) to thymine, which generates a G:T mismatch (Chatterjee and Walker 2017). If this mismatch escapes repair prior to DNA replication, it can result in a fixed C:G to T:A transition. In contrast, C:G to A:T mutations are often associated with oxidative damage to guanine, particularly the formation of 8-oxoguanine (Lynch et al. 2008). This lesion can mispair with adenine during DNA replication, leading to a G:C to T:A transversion, manifested as a C:G to A:T substitution.
Fig. 1
Mutation accumulation experiment in wild-type strains and mutants defective in non-homologous end joining (NHEJ) and translesion synthesis (TLS) pathways. A The wild type (WT) and mutants ku70, rev1, rev3, and rad30 were subcultured on YPD plates under UV- and MMS-treated conditions for multiple cycles. Nine or ten isolates for each strain were sequenced for mutation detection. B The spontaneous rates of SNVs and InDels of the wild type and mutants ku70, rev1, rev3, and rad30. The asterisk (*) indicates a significant difference from the WT at the 0.05 level (Mann–Whitney test). C Mutation spectrum (transitions and transversions) in the wild type and mutants ku70, rev1, rev3, and rad30
Among the detected InDels, 1-bp deletions were the most prevalent class, accounting for 56.8%, followed by 1-bp insertions at 28.6% (Dataset S1-2). Sequence context analysis revealed that over 90% of these InDels occurred within mononucleotide repeats or microsatellite regions, suggesting that replication slippage, rather than the NHEJ pathway, is the primary mechanism underlying these mutations. To test this hypothesis, we performed whole-genome sequencing on nine isolates derived from a NHEJ-deficient (ku70) mutant that had undergone 85 generations of subculturing. Our previous study demonstrated that deletion of KU70 in Y. lipolytica effectively inactivates the NHEJ pathway and prevents the majority of InDels induced by DNA-damaging agent Zeocin (Xiong et al. 2025). Here, the rate of InDels in these ku70 isolates subcultured under spontaneous conditions was estimated at 7.1 × 10−11 (28/9/25/85/20,500,000) per base per cell division, which was not significantly different from that observed in wild-type isolates (Mann–Whitney test, P > 0.05; Fig. 1B). These results confirm that the NHEJ pathway plays a minimal role in the generation of spontaneous small-scale InDels in Y. lipolytica.
To assess the role of the TLS pathway in spontaneous mutagenesis, we sequenced ten subcultured isolates from each Y. lipolytica mutant deficient in TLS polymerases: rev1, rev3, and rad30 (Xiong et al. 2025). We found that the deletion of REV1 (encoded by YALI1_F12757g, spanning base pairs 1,275,748–1,279,407 on chromosome F) and REV3 (encoded by YALI1_C29926g, spanning base pairs 2,992,626–2,996,642 on chromosome C) resulted in 50% and 40% reductions in SNV rates, respectively (Mann–Whitney test, P < 0.05; Fig. 1B). These results indicate that half of spontaneous SNVs are attributable to the activity of the TLS pathway involving Rev1 and DNA Pol ζ. In contrast, deletion of RAD30 (encoded by YALI1_C02152g, base pairs 213,307–215,229 on chromosome C), which encodes DNA Pol η, did not significantly affect the overall SNV rate. Furthermore, the relative proportions of specific base substitutions—including two transitions (C:G to T:A and T:A to C:G) and four transversions (C:G to T:A, A:T to C:G, C:G to G:C, and A:T to T:A)—remained unchanged in the absence of NHEJ or TLS polymerase activity under spontaneous conditions (Fisher’s exact test, P > 0.05, Fig. 1C).
In addition to point mutations, we identified three large duplications among the nine sequenced wild-type isolates: from 1630 to 2108 kb on chromosome E (Fig. 2A), 413 to 2251 kb on chromosome A (Fig. 2B), and 1209 to 1257 kb on chromosome C (Fig. 2C). The frequency of such large-scale rearrangements was estimated at 1.6 × 10−4 events per cell division. Similarly, three DNA fragment duplications were detected in the nine ku70 mutant isolates (Fig. 2D–F), indicating that the deficiency of the NHEJ pathway does not significantly impact the spontaneous rate of chromosomal rearrangement in Y. lipolytica. In the rad30, rev1, and rev3 mutants, we detected 2, 1, and 1 large duplications, respectively. However, the small number of observed events prevents us from drawing definitive conclusions regarding the potential suppression of large-scale rearrangements in these TLS-deficient mutants.
Fig. 2
Chromosomal rearrangements in the wild-type isolates and ku70mutant. Through calculating the sequencing coverage, three DNA duplications were identified in the isolates of wild type (A–C) and ku70 mutant (D–F), respectively. Yyl-1 and Yyl-3 are isolates of the wild-type Y. lipolytica strain, whereas YK3, YK9, and YK10 are isolates derived from the ku70 mutant background
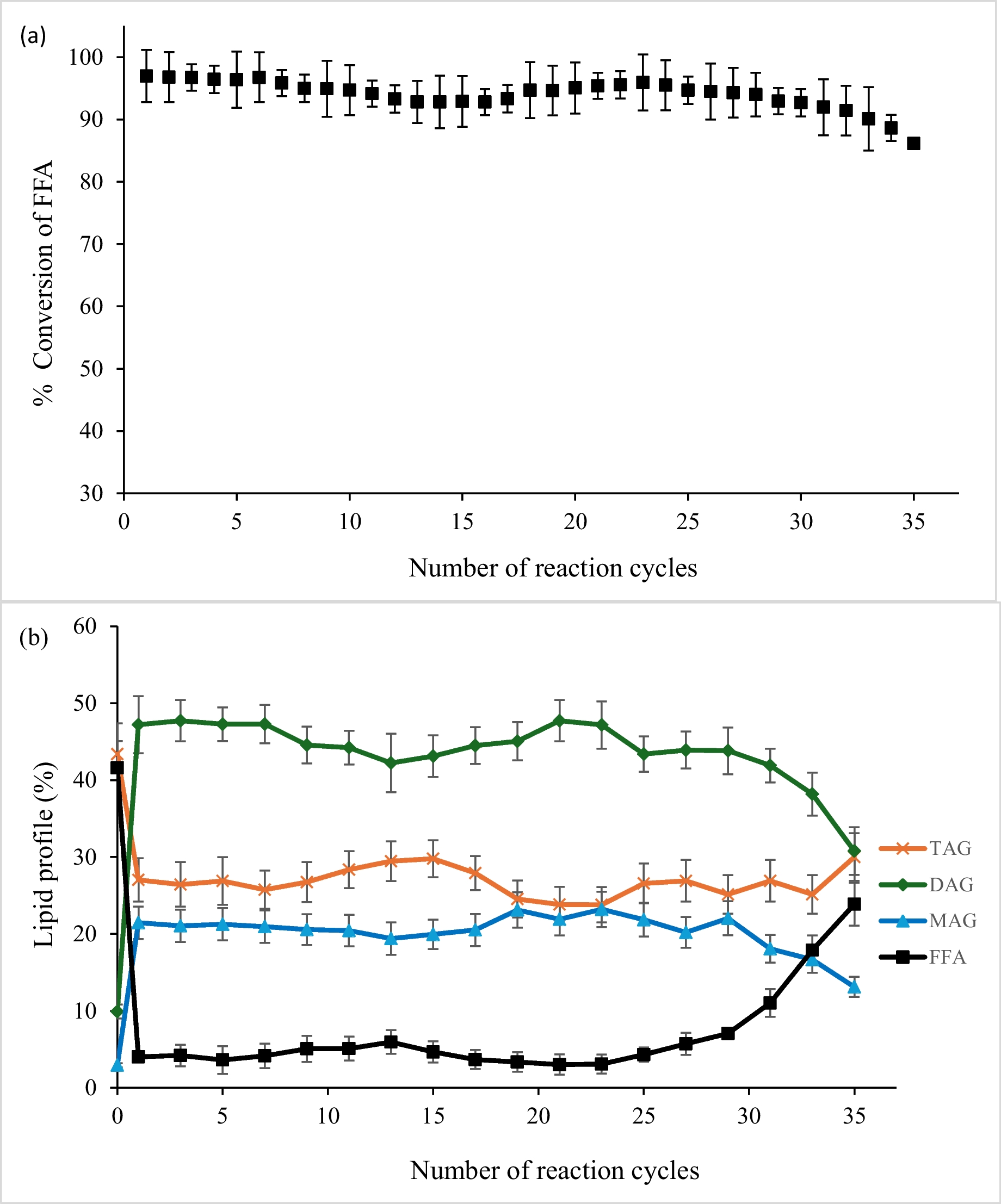
MMS-induced DNA mutations in wild-type strains and NHEJ- and TLS-deficient mutantsMMS is a well-known alkylating agent that introduces alkyl groups to DNA bases, particularly at the O6 and N7 positions of guanine (O6-MeG and N7-MeG) and the N3 position of adenine (N3-MeA), leading to the formation of unstable DNA adducts (Volkova et al. 2020). MMS has been widely used to study DNA repair mechanisms and for strain improvement in industrial applications (Dong et al. 2022). Under 0.02% MMS treatment, the ku70 and rad30 mutants exhibited biomass formation comparable to that of the wild-type strain after 24 h of incubation, while the rev1 and rev3 mutants displayed significantly slower growth (Fig. S1A in Supplemental file). To assess the mutagenesis of MMS in Y. lipolytica, we subcultured ten isolates of the wild-type strain PPF and NHEJ and TLS mutants on solid YPD containing 0.02% MMS for ten generations, followed by whole-genome sequencing. We identified 431 SNVs and 14 InDels among the ten isolates of wild-type strains treated with MMS, indicating their rates being 8.4 × 10−9 and 2.7 × 10−10 per base per cell division (Fig. 3A). These rates were 47- and threefold higher than that in the untreated isolates.
Fig. 3
MMS-induced mutations in wild-type isolates and mutants defective in NHEJ and TLS pathways. A The rates of SNVs and InDels of the wild-type strains and mutants rev1, rev3, and rad30 treated with MMS. The asterisk (*) denotes statistically significant differences (P < 0.05) compared to the WT, based on the Mann–Whitney U test. B Mutation spectrum in the wild-type strains and mutants rev1, rev3, and rad30. C Three-base motifs for spontaneous SNVs (top) and MMS-induced SNVs (bottom)
In ku70-deficient cells, the frequency of MMS-induced SNVs was reduced by approximately 40% relative to wild-type controls (Fig. 3A). A comparable decrease (37%) in SNV rate was observed upon deletion of RAD30, suggesting that both Ku70 and Rad30 contribute to the accumulation of MMS-induced mutations (Fig. 3A). Strikingly, the disruption of either REV1 or REV3 resulted in a dramatic reduction in MMS-induced mutagenesis (Fig. 3A). Specifically, the SNV rates in the rev1 and rev3 mutants were 7.2 × 10−10 and 8.8 × 10−10 per base per cell division, respectively—representing only 8.6% and 10% of the rate observed in MMS-treated wild-type strains (Fig. 3A). These results strongly support the conclusion that Rev1 and DNA Pol ζ constitute the primary pathway responsible for MMS-induced base substitution events.
In addition to increasing the overall mutation burden, MMS treatment induced a marked shift in the mutational spectrum. Specifically, A:T to T:A transversions became the predominant substitution type, comprising up to 56% of all SNVs following exposure (Fig. 3B). Analysis of the bases adjacent to MMS-induced A:T to T:A substitutions suggests a preference for G/C over A/T as the 5′ base of the mutated nucleotide (Fig. 3C; P < 0.05, Fisher’s exact test). This sequence context bias was not observed in spontaneous A:T to T:A substitutions (Fig. 3C). Additionally, the relative ratio of A:T to G:C was also significantly elevated by MMS treatment (Fig. 3B; P < 0.05, Fisher’s exact test). These striking enrichments represent a hallmark of MMS-induced DNA damage (particularly N3-MeA) and reflect its preferential resolution via error-prone repair pathways. Deletion of KU70 or RAD30 had no appreciable effect on the MMS-induced mutational profile. In contrast, loss of REV1 or REV3 effectively abolished this mutation signature, with the relative frequency of A:T to T:A transversions reduced to ~ 20% in both mutants (Figs. 3B and 4A). These findings underscore the essential role of Rev1 and DNA Pol ζ in shaping the mutational landscape associated with MMS-induced DNA lesions.
Fig. 4
The effect of REV1 and REV3 deletion on base substitutions in MMS-treated strains. A The rates of each individual base substitution in the wild-type strain and rev1 and rev3 mutants. B A model depicting the role of Rev1 and Pol ζ in the mutagenesis of MMS
Based on the above results, we propose a working model to illustrate the mechanism of MMS-induced mutagenesis in Y. lipolytica, as depicted in Fig. 4B. In this model, O6-MeG and N3-MeA are considered the primary mutagenic lesions. O6-MeG is a highly mutagenic adduct that mispairs with thymine during DNA replication instead of its canonical partner cytosine. If this mispair escapes repair prior to the next round of replication, it results in G:C to A:T transition mutations. Given that deletion of REV1 and REV3 only modestly reduced the frequency of C:G to T:A mutations, it is likely that the insertion of thymine opposite O6-MeG is predominantly carried out by the replicative DNA Pol δ and Pol ε, rather than by the TLS pathway. In contrast, N3-MeA is a replication-blocking lesion that interferes with base pairing by disrupting the N3 position of the adenine ring—an essential site for hydrogen bonding in the DNA duplex. This damage stalls the progression of replicative polymerases δ and ε. To bypass this blockage, Y. lipolytica cells recruit the TLS polymerases Rev1 and Pol ζ, which insert nucleotides in an error-prone manner. Our data suggest that adenine is the most frequently inserted nucleotide opposite N3-MeA, consistent with the predominance of A to T transversions observed in MMS-treated cells.
The roles of TLS and NHEJ pathways in UV-induced mutations in Y. lipolyticaUV radiation can directly damage DNA by inducing the formation of photoproducts, such as cyclobutane pyrimidine dimers (CPDs) and 6–4 photoproducts (6-4PPs), which disrupt the normal structure of the DNA helix (de Lima-Bessa et al. 2008). To investigate the mutagenic effects of UV exposure on Y. lipolytica, ten isolates derived from wild-type, NHEJ-deficient, and TLS-deficient strains were subjected to ten cycles of UV treatment at a dose of 80 J/m2 (Fig. 1A). At this UV dosage, the viability of wild-type Y. lipolytica strain PPF was approximately 86% (Fig. S1B). In contrast, the ku70, rad30, rev1, and rev3 mutants exhibited markedly reduced viabilities of 49%, 34%, 16%, and 15%, respectively (Fig. S1B). In UV-treated wild-type isolates, we identified a total of 964 SNVs and 18 InDels (Datasets S1 and S2). Compared to spontaneous conditions (Fig. 1B), UV exposure resulted in a 107-fold increase in the SNV rate (1.9 × 10−8 per base per cell division) and a 3.7-fold increase in the InDels rate (3.5 × 10−10 per base per cell division) (Fig. 5A). These findings suggest that UV radiation predominantly induces base substitutions rather than small InDels in Y. lipolytica, consistent with our recent study conducted in a different genetic background (Xiong et al. 2025). Notably, deletion of REV1 and REV3 significantly attenuated UV-induced mutagenesis, resulting in 85% and 80% reductions in SNV rates, respectively (Mann–Whitney test, P < 0.05; Fig. 5A). These results demonstrate that the majority of UV-induced SNVs are dependent on the TLS polymerases Rev1 and Pol ζ. In contrast, ku70 and rad30 mutants exhibited 1.3- and 1.4-fold higher SNV rates, respectively, compared to the wild-type strains (Fig. 5A), indicating that both Ku70 and Pol η (Rad30) contribute to the suppression of UV-induced mutations.
Fig. 5
UV-induced mutations in wild-type strains and mutants defective in NHEJ and TLS pathways. A The rates of SNVs and InDels of the wild-type strains and mutants rev1, rev3, and rad30 treated with UV. The asterisk (*) indicates a significant difference from the WT at the 0.05 level, as determined by the Mann–Whitney U test. B Mutation spectrum in the wild-type strains and mutants rev1, rev3, and rad30. C Three-base motifs for UV-induced SNVs
Under spontaneous conditions, A:T to G:C substitutions accounted for only 8.7% of SNVs (Fig. 5B). Upon UV treatment, however, the frequency of this substitution type increased significantly to 24% in the wild-type Y. lipolytica strain (P < 0.05, Fisher’s exact test), representing the most strongly elevated base change. UV exposure also markedly increased the proportion of C:G to T:A substitutions, which rose to 56% (P < 0.05, Fisher’s exact test; Fig. 5B). Notably, this UV-induced increase in C:G to T:A substitutions was further amplified in mutants lacking RAD30, REV1, or REV3, indicating that these TLS polymerases play a key role in modulating the mutational spectrum under UV stress. In contrast, deletion of KU70 had no significant effect on either the SNV spectrum or InDels frequency (Fig. 5A, B).
Pol η avoids C to T and C to A mutations but contributes to T to C in UV-treated Y. lipolyticaAlthough the deletion of RAD30 moderately increased the overall SNV rate in UV-treated Y. lipolytica cells, its effects on specific base substitution types were distinct. Notably, the rates of C:G to T:A and C:G to A:T substitutions were elevated by 1.7- and 4.4-fold, respectively, in the rad30 mutant compared to that in the wild-type strain (Mann–Whitney test, P > 0.05; Fig. 6A). In contrast, the frequency of A:T to G:C substitutions was reduced by approximately 65% (Mann–Whitney test, P > 0.05; Fig. 6A). These findings underscore the dual role of DNA Pol η in the UV-induced mutagenic response of Y. lipolytica—acting both to suppress and to generate specific mutation types depending on the lesion context.
Fig. 6
The effect of RAD30 deletion on base substitutions in UV-treated strains. A The rates of each individual base substitution in the wild-type strain and rad30 mutant. The asterisk (*) indicates a significant difference at the 0.05 level compared to WT. B A model depicting the role of Rev1, Pol ζ, and Pol η in the mutagenesis of UV in Y. lipolytica
UV irradiation predominantly induces CPDs, particularly at dipyrimidine sites such as TT, TC, CT, and CC (Premi et al. 2015). C:G to T:A and C:G to A:T substitutions are generally associated with cytosine-containing CPDs (TC or CC), where cytosine deamination or mispairing may occur during error-prone bypass in the absence of Pol η (Washington et al. 2001). Analysis of the adjacent base of SNVs confirmed that most mutations in UV-treated Y. lipolytica cells were associated with CPDs (Fig. 5C). The increased frequency of these mutations in the rad30 mutant suggests that alternative translesion polymerases, such as Rev1 or Pol ζ, may bypass these lesions in a more error-prone manner when Pol η is absent.
In Fig. 5C, we observed that the majority of T to C mutations occurred within 5′-TT-3′ or 5′-CT-3′ sequence contexts. This pattern suggests that these base substitutions may be associated with CPDs or 6-4PPs. Given that previous studies have shown that DNA Pol η in S. cerevisiae can introduce T to C substitutions during replication across site-specific 6-4PPs (Bresson and Fuchs 2002), we propose that the T to C substitutions observed in UV-treated Y. lipolytica are likewise the result of error-prone bypass of 6-4PPs mediated by Pol η (Fig. 6B). Together, these observations highlight the lesion-type-specific roles of Pol η in Y. lipolytica, balancing accurate translesion synthesis and mutagenesis under UV-induced DNA damage.
Evolved resistance to oxidative stress in Y. lipolytica requires NHEJ and TLSOur above analysis confirmed the distinct contributions of NHEJ and TLS polymerases to spontaneous and genotoxic stress–induced DNA mutations. We then considered whether decreasing the activity of NHEJ and TLS in cells would affect the kinetics of phenotypic evolution. To test this hypothesis, we utilized a laboratory evolution assay as described in the “Materials and methods” section. During this assay, we measured the adaptation to two oxidative stressors, Zeocin and H2O2, in the wild-type and ku70, rev1, rev3, and rad30 mutants. Although all tested strains showed gradually increased MIC50 values during the 7-day evolution, we observed that the wild-type strain PPF and rad30 mutants evolved more rapidly than the ku70, rev1, and rev3 mutants in Zeocin-containing medium (Fig. 7A). Zeocin, a member of the bleomycin antibiotic family, binds directly to DNA and induces both strand breaks and oxidative base damage (Chen and Stubbe 2005; Zheng et al. 2022). Our previous study demonstrated that Ku70 and DNA Pol ζ are responsible for approximately 80% of Zeocin-induced InDels and 67% of SNVs in Y. lipolytica (Xiong et al. 2025). These findings suggest that the reduced mutagenic capacity of NHEJ- and TLS-deficient mutants may impair their ability to generate adaptive genetic variations under Zeocin-induced stress, thereby limiting their evolutionary potential. Similarly, under H2O2 stress, rev1, rev3, and ku70 mutants exhibited a lower tendency to evolve increased resistance compared to the wild-type strain and rad30 mutant (Fig. 7B). These results demonstrated the contribution of NHEJ and TLS in the phenotypic evolution of Y. lipolytica under oxidative stress.
Fig. 7
Resistance evolution of Y. lipolytica strains under Zeocin and H2O2stress. MIC50 values of the wild-type strain PPF and the mutants rev1, rev3, and rad30 during the continuous passages in the presence of A Zeocin and B H2O2
Comments (0)