
Remember me

In m-IHC Cohort, we included baseline pre-treatment samples from 172 patients, which were divided into four panels for staining (Fig. 1a, b). The median age of the patients was 62.00 years (range, 53.00–69.00), with 75.6% of the patients being male (Table 1). Eighteen (10.5%) patients had EBV-positive, and twelve (7.0%) had confirmed deficient DNA mismatch repair (dMMR) GC. We calculated the VISTA expression density in the total region (tumour + stroma) for each sample, defining the top 30% as high expression and the bottom 70% as low expression (cut-off value: 130 cell/mm²), and we conducted sensitivity analysis using different VISTA expression cut-offs to validate the robustness of our findings (Supplementary Fig. S3). Additionally, samples were categorised based on VISTA expression levels in the tumour and stromal regions (Supplementary Table S1, 2). Comparison between the tumour and stromal regions showed no difference in overall VISTA expression (Fig. 2a). However, the density of VISTA+ M2 macrophages (VISTA+ CD68+ CD163+ CD86-) was significantly higher in the tumour region than in the stromal region (P = 0.017) (Fig. 2a).
Fig. 1: Identification and characterisation of the tumour microenvironment in GC patients according to VISTA expression.
A Study flow chart and analysis design of the immunohistochemistry cohort. B The merged and single-stained images for 4 representative panels of multiplex IHC. Scale bar: 50 μm. C Study flow chart of single-cell RNA sequencing cohort. D Study flow chart of the spatial transcriptomics cohort. Created in BioRender.
Fig. 2: The association of VISTA expression with clinicopathological and prognostic features in GC.
a The comparison of VISTA expression and VISTA+ M2-like macrophage density in Tumour/Stroma regions. Mann–Whitney U test. ns: not significant. B The association between VISTA expression in the total region and clinicopathological features. Mann–Whitney U test. ns: not significant. C Overall survival, immunotherapy related OS (irOS), immunotherapy related PFS (irPFS) of patients based on VISTA expression in total region and tumour region. The irOS and irPFS were calculated within immunotherapy-population. Log-rank (Mantel-Cox) test. HR: hazard ratio. HRs with 95% confidence interval (95% CI) and corresponding P-values were adjusted for the following covariates: age, ECOG performance status, tumour location, histological differentiation grade, Lauren classification, disease stage, HER2 expression status, PD-L1 expression level, MMR status, EBV status, and line of therapy.
Table 1 The clinicopathological features between VISTA-high group and VISTA-low group based on Total regions.The expression level of VISTA in stage III samples was significantly higher than in stage IV samples (Fig. 2b). However, no association was found between VISTA expression and age, gender, ECOG performance status, tumour location, tumour differentiation status, Lauren classification, HER2 status, PD-L1 status, or MMR status (Fig. 2b). Furthermore, we analysed the prognostic value of VISTA expression. There was no significant association between high VISTA expression in the total or stromal regions and prognosis (Fig. 2c, and Supplementary Fig. S1c). However, high VISTA expression in the tumour region was associated with poorer irPFS in the population receiving immunotherapy (HR = 3.367 (95% CI: 1.605–7.064), P = 0.001) (Fig. 2c).
VISTA high expression correlates with a suppressive immune microenvironmentWe analysed the differences in the immune microenvironment of GC samples with high VISTA expression, focusing on the comparison of expression differences in CD8+ T cells, CD4+ T cells, fibroblasts, macrophages, neutrophils, and various immune checkpoints.
In the total region, no significant differences were observed in the density of non-exhausted CD8+ T cells between VISTA-high group and low group. However, a significant enrichment of CD8+ TIGIT+ associated T cell subsets (including CD8+ PD-1- TIGIT+ T cells and CD8+ TIGIT+ TIM3- T cells) was observed (Fig. 3a, b). Meanwhile, in the VISTA-high group, there was a significant increase in the expression of CD4+ cells, including both effector CD4+ T cells (including CD4+ FOXP3-, CD4+ FOXP3- PD-L1-, and CD4+ FOXP3- CTLA4-) and regulatory T cells (Treg cells, including CD4+ FOXP3+, CD4+ FOXP3+ PD-L1+, CD4+ CTLA4+ FOXP3+, and CD4+ CTLA4+ FOXP3+ PD-L1+) (Fig. 3a, b). In addition, M1-like macrophages (CD68+ CD163- CD86+), M2-like macrophages (CD68+ CD163+ CD86-), fibroblasts (FAP+), neutrophils (CD66B+), and several immune checkpoints (including PD-L1 and TIGIT) were enriched in the VISTA-high group (Fig. 3a, b).
Fig. 3: Expression patterns of unique TIICs in VISTA-high/low tissues.
a, b The difference analysis of TIIC density between VISTA-high tissues and VISTA-low tissues. Significant: P < 0.05. Not Significant: P > 0.05. c Expression correlation between VISTA+ cells and other unique TIICs in the total region. ns: not significant. d Density of M1/2-like macrophages within total region in GC. e, f Comparison of the density of detailed macrophage subtypes among VISTA-positive macrophages in the total region. ns: not significant.
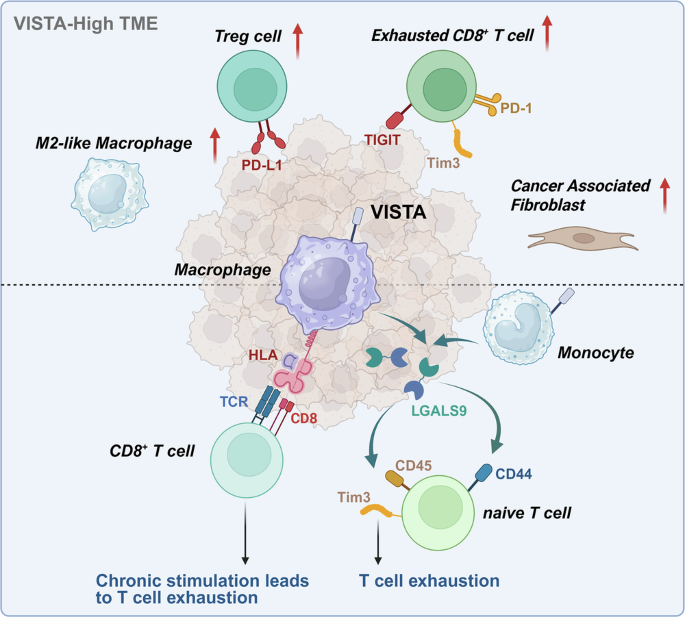
We further performed Pearson correlation analyses between VISTA expression and various immune checkpoints/cellular subpopulations in total, tumour and stroma regions (Fig. 3c, and Supplementary Fig. S4a, b). VISTA demonstrated strong positive correlations with VISTA⁺ macrophages (CD68⁺ VISTA⁺, r = 0.98) in total region and VISTA⁺ tumour cells in tumour region (VISTA⁺ PANCK⁺, r = 0.98) (Fig. 3c, and Supplementary Fig. S4a). Moreover, VISTA expression showed positive associations with FOXP3+ Treg cells (CD4+ FOXP3+ , r = 0.35, P < 0.001) and neutrophil (CD66B+, r = 0.64, P < 0.001) abundance, but not with FOXP3- effector CD4+ T cells (CD4+ FOXP3-, P > 0.05) (Fig. 3c). Among immune checkpoints, PD-L1 exhibited the strongest correlation with VISTA expression (Total region: r = 0.35; Tumour region: r = 0.29; Stroma region: r = 0.47) (Fig. 3c, and Supplementary Fig. S4a, b). In summary, samples with high VISTA expression exhibited an inhibiting immune phenotype, where immune cells exerted immunosuppressive effects (such as exhausted CD8+ T cells, Treg cells, and M2 macrophages) in the VISTA-high GC TME.
Previous studies indicate predominant VISTA expression in myeloid cells within the TME, particularly macrophages [35]. We therefore quantified VISTA⁺ cell proportions across macrophage subtypes using m-IHC. While notable VISTA⁺ M1-like macrophage infiltration was observed in select specimens (Fig. 3d), comparative analysis revealed no statistically significant difference in cell densities between VISTA⁺ M1-like and VISTA⁺ M2-like macrophage populations (Fig. 3e). Given the emerging role of stimulator of interferon genes (STING) as a macrophage-associated pro-tumorigenic/immunosuppressive protein in GC, we evaluated STING-associated macrophage subsets [36]. Quantitative assessment demonstrated significantly reduced infiltration densities in VISTA⁺ M1 STING⁻ macrophages compared to other subsets (VISTA⁺ M1 STING⁺, VISTA⁺ M2 STING⁺, and VISTA⁺ M2 STING⁻). No significant inter-group differences were detected among the latter three subpopulations.
Inhibiting spatial immune cell features in VISTA-High GC samplesWe used the developed spatial analysis tool to assess the spatial features in the VISTA-high group, focusing on differential analysis of the Effective score and Effective percent (Fig. 4a, b). Effective percent and Effective score calculate the densities and proportions of multiple immune cell types within a 20 μm radius of tumour cells (see Methods) [14].
Fig. 4: Spatial distribution of TIICs in VISTA-High GC.
a Graphical depiction of spatial relationships between tumour and immune cells. Tumour cells are shown as green dots, immune cells as red dots, and connections via white lines indicate distances of less than 20 microns. b Differences in effective score were analysed between VISTA-high and VISTA-low groups. c Differences in effective percent were analysed between VISTA-high and VISTA-low groups. Mann–Whitney U test. d Kaplan-Meier survival analysis of VISTA+ M2-like macrophages stratified by effective score (high score defined as ≥70th percentile). Log-rank (Mantel-Cox) test. HR: hazard ratio. HRs with 95% confidence interval (95% CI) and corresponding P-values were adjusted for the following covariates: age, ECOG performance status, tumour location, histological differentiation grade, Lauren classification, disease stage, HER2 expression status, PD-L1 expression, MMR status, EBV status, and line of therapy. a Created in BioRender.
The VISTA-high group exhibited significantly elevated effective score across multiple immune subsets compared to low group (Fig. 4b, and Supplementary Fig. S5a, b, Supplementary Table S4). Specifically, we observed marked increases in exhausted T cells (CD8⁺ TIGIT⁺: 0.038 vs. 0.012, P = 0.003), Treg cells (CD4⁺ FOXP3⁺: 0.042 vs. 0.011, P < 0.001), M1-like macrophages (CD163⁻ CD68⁺ CD86⁺ STING⁺: 0.055 vs. 0.010, P < 0.001), M2-like macrophages (CD163⁺ CD68⁺ CD86⁻ STING⁺: 0.092 vs. 0.026, P < 0.001), neutrophils (CD66b⁺: 0.758 vs. 0.299, P < 0.001), and fibroblasts (FAP⁺: 1.929 vs. 1.509, P = 0.001). Effective percent measurements demonstrated comparable patterns to absolute efficiency scores (Fig. 4c, and Supplementary Fig. S5b; Supplementary Table S5). In VISTA-high group, significant elevations of effective percent were observed across multiple immune subsets: exhausted T cells (CD8⁺ PD1⁺: 0.051 vs. 0.021, P = 0.018; CD8⁺ TIGIT⁺: 0.030 vs. 0.010, P = 0.003), Treg cells (CD4⁺ FOXP3⁺: 0.036 vs. 0.010, P < 0.001), M1-like macrophages (CD163⁻ CD68⁺ CD86⁺ STING⁺: 0.041 vs. 0.009, P < 0.001), M2-like macrophages (CD163⁺ CD68⁺ CD86⁻ STING⁺: 0.067 vs. 0.022, P < 0.001), neutrophils (CD66b⁺: 0.336 vs. 0.175, P < 0.001), and fibroblasts (FAP⁺: 0.638 vs. 0.567, P = 0.009).
We also investigated the relationship between the Effective score and the Effective percent of VISTA+ macrophages and the prognosis of GC patients. Patients were categorised into high and low groups based on the 70th percentile of the effective score and effective percent. Our results indicate that higher effective score/percent of VISTA+ macrophages correlate with worse irPFS in gastric cancer patients (score: HR = 3.036 (95% CI: 1.478 - 6.195), P = 0.002; percent: HR = 2.879 (95% CI: 1.409–5.881), P = 0.002), but no statistical association was found with OS/irOS (Supplementary Figs. S6a, b). We separately assessed the relationship between the Effective Score/percent of VISTA+ M1-like and VISTA+ M2-like macrophages and prognosis. The data showed that higher Effective score of VISTA+ M2-like macrophages was associated with worse OS (HR = 1.812 (95% CI: 1.016–3.232), P = 0.044), irOS (HR = 2.053 (95% CI: 1.111–3.794), P = 0.022), and irPFS (HR = 2.141 (95% CI: 1.069–4.288), P = 0.032) (Fig. 4d), while no significant association was observed for VISTA+ M1-like macrophages (Supplementary Figs. S6C, D).
Single-cell RNA sequencing analysis of VSIR expression in specific monocyte-macrophage subsets in gastric cancerWe performed scRNA-seq analysis on 17 GC samples collected from scRNA-seq Cohort. After quality control, we obtained 108,033 high-quality cells (Supplementary Fig. S7A). The cells were clustered into 27 subpopulations and annotated into 10 distinct subtypes: T & NK (Nature Killer) cells, plasma cells, B cells, myeloid cells, mast cells, fibroblasts, epithelial cells, endothelial cells, endocrine cells, and smooth muscle cells (Fig. 5A). The ROGUE scores of most cell subpopulations exceeded 0.7, while the ROGUE score for myeloid cells was relatively lower, suggesting the potential for further subdivision of the myeloid compartment (Supplementary Fig. S7C). We found that VSIR, the gene encoding VISTA, was prominently expressed in myeloid cells (Fig. 5C, D).
Fig. 5: Single-cell transcriptomic atlas of GC patients reveals the expression landscape of VSIR across distinct cell populations.
a Single-cell transcriptomic atlas of GC patients, showing distinct cell clusters including T&NK cells, B cells, plasma cells, myeloid cells, mast cells, endocrine cells, epithelial cells, endothelial cells, fibroblasts, and smooth muscle cells. b UMAP dimensionality reduction plots of myeloid cells subpopulations and the classification of various subsets. c VSIR expression mapped onto UMAP. Blue circles: high VSIR expression in myeloid cells. d The expression levels of VSIR across different cellular subpopulations were compared, revealing that its expression in myeloid cells was significantly higher than in other subpopulations. e Expression levels of VSIR in different myeloid cell populations, with the median values annotated in the bar chart. F Different subsets of Monocyte-Macrophage cells exhibit distinct characteristic M1/M2/antigen presentation functional genes. g Functional scores of multiple gene sets in monocyte-macrophage populations, calculated using UCell. h Comparison of gene set functional scores between VSIR+ and VSIR- cells in monocyte-macrophage subpopulations. *P < 0.05, **P < 0.01, ***P < 0.001.
To further characterise the expression pattern of VSIR, we performed a finer subdivision of each cell category, with a particular focus on the myeloid compartment, which was further classified into macrophages, monocytes, dendritic cells (DCs), and neutrophils (Fig. 5b, and Supplementary Fig. S9a). The differential gene expression across subpopulations of myeloid cells is shown in Fig. 5e. We found that VSIR was primarily expressed in monocyte-macrophage (MoMac) subpopulations, including C3+ macrophages (median expression value: 1.28), FCN1+ monocytes (median expression value: 1.11), CD163L1+ macrophages (median expression value: 0.97), PLK1+ macrophages (median expression value: 0.77), SPP1+ macrophages (median expression value: 0.67), but not in DCs or neutrophils (Fig. 5e). We further scored the myeloid cells to assess the functional characteristics of various cell types within the MoMac subpopulation (Fig. 5g). The results revealed that VSIR was highly expressed in MoMacs with enhanced antigen presentation functions (C3+ macrophages and CD163L1+ macrophages) and M2-like macrophages (CD163L1+ macrophages and SPP1+ macrophages), rather than in the typical M1 macrophage subtypes (M1-like CXCL5+ macrophages) (Fig. 5f, g). We compared various functional scores between VSIR+ and VSIR- cells within each MoMac subpopulation due to the expression pattern (Supplementary Fig. S10c). The results showed that VSIR+ cells in each subpopulation exhibited higher antigen presentation score(Fig. 5h, Supplementary Table S8). However, no significant differences were observed in the M1 score within the VSIR high-expression subpopulations (C3+ macrophages: 0.1311 vs. 0.1402, P = 0.528; FCN1+ monocytes: 0.1516 vs. 0.1484, P = 0.104). In conclusion, these findings indicate that VSIR is expressed in macrophages with enhanced antigen presentation functions in GC TME, rather than in the M1 macrophage subtypes.
VSIR+ MoMac cells promote T cell exhaustion through the LGALS9-PTPRC pathway and antigen presentationWe performed differentiation trajectory analysis of MoMac cells using the Monocle and Slingshot packages, which yielded similar results (Fig. 6a, and Supplementary Fig. S11a). The differentiation trajectory revealed how MoMac cells differentiate from FCN1+ monocytes into various macrophage subpopulations, with a progressive shift toward the M2-like phenotype. During this process, antigen presentation and phagocytosis gradually increased (Fig. 6d). VSIR was prominently expressed in early-stage FCN1+ monocytes, early-to-mid-stage C3+ macrophages and late-stage CD163L1+ macrophages (Fig. 6b–e).
Fig. 6: Pseudotime Analysis and Cell communications Analysis of VSIR+ monocyte-macrophages in GC TME.
a Pseudotime trajectory depicting the dynamics of monocyte-macrophages (MoMacs). b Distribution of VSIR along the pseudotime trajectory in MoMacs. c Distribution of multiple MoMac cell subtypes along the pseudotime trajectory. d Dynamics of gene set functional scores along the pseudotime trajectory. e Identification of VSIR along the pseudotime trajectory. f, g Cell communication patterns upregulated and downregulated in VISR+ MoMacs compared to VSIR- MoMacs with other cell subsets and various T cell subpopulations. Red indicates a higher probability of communication in VISR+ MoMacs, while blue indicates a lower probability. h Different subsets of T cells exhibit distinct characteristic naïve/exhausted/cytotoxcity functional genes. i Functional scores of multiple gene sets in T cell populations, calculated using UCell package. J. Spatial co-localisation of C3+ macrophages and T cells in ST1. k Inference of LGALS9-CD45 pathway distribution on spatial sections. l, m Differentially expressed genes (DEGs) and immune-related pathway enrichment analysis in co-localised regions of C3+ macrophages and T cells.
Additionally, we described the differences in cell communication between VSIR+ and VSIR- MoMac cells. We found that VSIR+ MoMac cells predominantly interact with T & NK cells through the LGALS9 (Galectin-9)–PTPRC (CD45)/CD44 and HLA-CD8 pathways, and with B cells through the APP-CD74 pathway (Fig. 6f–h). We further examined the interactions between VSIR+ MoMac and T/B lymphocytes, scoring various T cell functions and performing temporal analysis to describe their roles (Fig. 6h, i, and Supplementary Fig. S12a–h). The results revealed that, compared to VSIR- MoMac cells, VSIR+ MoMac cells preferentially interact with early-stage T cells (CD8+ GZMK+ Tem, CD4+ ICA1+ Treg, and CD4+ LEF1+ Tn) through the LGALS9–PTPRC/CD44 pathway (Fig. 6h, i). VSIR+ MoMacs also continuously stimulate CD8+ GZMK+ Tem and CD8+ ZNF683+ Tm cells through antigen presentation (Fig. 6G). The latter highly express PDCD1, LAG3, and HAVCR2, exhibiting characteristics of exhausted CD8+ T cells. This suggests that VSIR+ MoMacs may promote the differentiation of early CD8+ GZMK+ Tem into exhausted T cells through chronic antigen presentation [37]. Additionally, we stratified the samples based on the expression levels of VSIR in MoMac cells and found that in VSIR-high samples, the numbers of T cells, particularly CD4+ Treg cells and exhausted CD8+ T cells, were increased, which further demonstrated the role of VSIR in promoting T cell exhaustion (Supplementary Fig. S13c).
We further aimed to characterise VSIR-related expression and functional features using ST. In all three ST samples, VSIR exhibited a distinct nest-like spatial clustering pattern. C3+ macrophages spatially co-localised with T cells, suggesting potential interactions between the two cell types. Notably, the LGALS9-PTPRC interaction between C3+ macrophages and T cells was also validated in the ST data. Enrichment analysis revealed that C3+ macrophages were involved in both antigen presentation and immunosuppression. Interestingly, the dominance of these functions varied across samples: ST1 predominantly exhibited immunosuppression (Fig. 6q), ST3 showed primarily antigen presentation activity (Supplementary Fig. S14r), and ST2 displayed a balanced profile between the two (Supplementary Fig. S14k).
Overall, VSIR+ MoMac cells exhibit enhanced antigen presentation, which induced T cell exhaustion. And through LGALS9-related and APP-related pathways, they suppress the activation of early T cells and the functionality of cytotoxic T cells.
Comments (0)