
Remember me
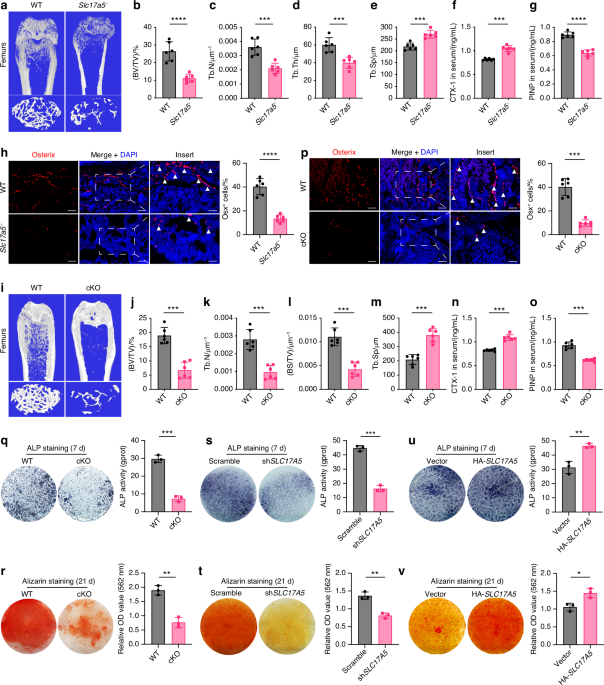
We constructed Slc17a5−/− mice and found that Sialin plays a key role in maintaining bone homeostasis in mice. Western blot analysis indicated successful deletion of Slc17a5 (Fig. S1a). Micro-computed tomography (μCT) images of the femur are shown in Fig. 1a. The results of bone histomorphometry indexes were obtained through quantitative analysis. The bone volume fraction (BV/TV), trabecular thickness (Tb.Th), and number of trabeculae (Tb.N) of the femur in Slc17a5−/− mice were lower than those in the wild-type (WT) control group (Fig. 1b–d), while the trabecular separation (Tb.Sp) was higher in Slc17a5−/− mice than that in the control group (Fig. 1e), indicating that Slc17a5−/− mice had relatively low bone mass. At the femoral mid-diaphysis, cortical bone volume (Ct.BV), cortical thickness (Ct.Th), and total trabecular area (Tt.Ar) were markedly decreased in Slc17a5−/− mice, consistent with the trabecular phenotype (Fig. S1d–f). The bone resorption marker collagen type I cross-linked C-telopeptide (CTX-1) were significantly elevated in Slc17a5−/− mice compared to that in WT (Fig. 1f). The bone formation marker procollagen type I N-propeptide (P1NP) was significantly decreased in Slc17a5−/− mice compared to that in WT (Fig. 1g). These results suggest that Sialin loss impairs bone homeostasis by altering bone remodeling. Immunofluorescence (IF) staining of bone sections revealed lower number of Osterix (Osx)-positive osteoblasts in Slc17a5−/− mice than in the control mice (Fig. 1h). We further performed tartrate-resistant acid phosphatase (TRAP) staining on femoral sections and found that Slc17a5−/− mice exhibited increased osteoclast formation compared to control mice (Fig. S1b).
Fig. 1
Sialin promotes osteogenic differentiation of mesenchymal stem cells and maintains bone homeostasis. a Representative micro-CT images of femurs from WT and Slc17a5-/- mice. Upper scale bars, 1 mm. Lower scale bars, 200 μm. Quantification of bone volume/tissue volume (BV/TV, b), trabecular number (Tb.N, c), trabecular thickness (Tb.Th, d), and trabecular separation (Tb.Sp, e) in WT and Slc17a5-/- mice. Serum levels of CTX-1 (f) and PINP (g) in WT and Slc17a5-/- mice. h Immunofluorescence (IF) staining images (left) and quantification (right) of osterix (Osx) in femoral sections from WT and Slc17a5-/- mice. Scale bars, 50 μm. Arrowheads indicate Osx-positive osteoblasts. i Representative micro-CT images of femurs from WT and cKO mice. Upper scale bars, 1 mm. Lower scale bars, 200 μm. Quantification of BV/TV (j), Tb. N (k), bone surface/tissue volume (BS/TV, l), and Tb.Sp (m) in WT and cKO mice. Serum CTX-1 (n) and PINP (o) levels in WT and cKO mice. p IF staining images (left) and quantification (right) of Osx in femoral sections from WT and cKO mice. Scale bars, 50 μm. Arrowheads indicate Osx-positive osteoblasts. ALP staining and activity (q), and ARS staining with quantification of mineralized nodules (r) in murine MSCs (mMSCs) from 8-week-old WT and cKO male mice. ALP staining and activity (s), and ARS staining and mineralization (t) in control and SLC17A5 knockdown (shSLC17A5) human MSCs (hMSCs). ALP staining and activity (u), and ARS staining and mineralization (v) in control and Sialin-overexpressing (HA-SLC17A5) hMSCs. Data are presented as the mean ± SD, n = 3, except for a–p (n = 6). *P < 0.05; **P < 0.01; ***P < 0.001
To verify the effect of Slc17a5 expression in MSCs on bone formation, we generated Slc17a5 flox (Slc17a5fl/fl) mice using CRISPR/Cas9 technique and bred Slc17a5fl/fl mice with Prx1-Cre transgenic mice to obtain conditional homozygous Slc17a5 knockout (KO), Prx1-Cre; Slc17a5fl/fl (cKO) mice. Western blot analysis indicated successful deletion of Slc17a5 in stromal cells isolated from long bones of Slc17a5 knockout mice (Fig. 1s). Micro-CT images of the femur are shown in Fig. 1i. The BV/TV, Tb.N, and bone surface fraction (BS/BV) of the femur in cKO mice were lower than those in the control group (Fig. 1j–l), while the Tb.Sp was in cKO mice higher than that in the control group (Fig. 1m), indicating that SLC17A5 may regulate osteogenic differentiation in MSCs and bone formation in mice. Similarly, in cKO mice, cortical bone indices were significantly reduced compared with their littermate WT controls. Ct.BV, Ct.Th, and Tt.Ar all showed decreases, again paralleling the trabecular findings (Fig. S1g–i). CTX-1 level was significantly elevated (Fig. 1n), while P1NP level was significantly decreased (Fig. 1o) in cKO mice compared to those in the WT. IF staining of bone sections revealed a lower number of Osx+ osteoblasts in cKO mice than in the WT (Fig. 1p). TRAP staining of femoral sections indicated elevated numbers of TRAP+ osteoclasts in cKO mice relative to controls (Fig. S1c). To verify whether Sialin regulates bone homeostasis by modulating the function of MSCs, we isolated MSCs from cKO and control mice. Significant reductions in alkaline phosphatase (ALP) staining intensity and activity, and alizarin red S (ARS) staining intensity and generation of mineralized nodules were noticed in MSCs of cKO mice on days 7 and 21 of differentiation, respectively (Fig. 1q, r). The mRNA expression of the key osteogenesis-related genes, such as Alpl, Runx2, and Spp1 (Fig. S1j–l), and expression of osteogenesis-related proteins, such as COL-I, RUNX2, and OCN (Fig. S1m), were also significantly downregulated, confirming the effect of Sialin on the osteogenic potential of murine MSCs.
To further clarify whether Sialin affects cellular osteogenic differentiation, we knocked down SLC17A5(Slc17a5) in human MSCs (hMSCs) and murine MSCs (mMSCs) using short hairpin (sh) RNA sequences. Sialin expression was significantly decreased (Figs. 1v and S2a). After 7 days of differentiation, the ALP staining intensity of SLC17A5(Slc17a5) knockdown hMSCs and mMSCs was markedly reduced compared to their respective controls, with a similar trend observed in the ALP activity assays (Figs. 1t and S2b). After 21 days of differentiation, ARS staining and quantitative analysis showed that the formation of mineralized nodules was significantly reduced in SLC17A5(Slc17a5) knockdown hMSCs and mMSCs (Figs. 1u and S2c). In hMSCs, the mRNA expression of osteogenesis-related genes, including ALPL, COL1A1, BGLAP, RUNX2, and SP7 (Fig. S1n–r), and the protein expression of COL-I, RUNX2, and OCN were markedly decreased (Fig. S1s).
Next, we overexpressed SLC17A5(Slc17a5) in hMSCs and mMSCs, which led to a significant increase in Sialin protein levels (Figs. 1y and S2d). After 7 days of differentiation, the ALP staining intensities of SLC17A5(Slc17a5)-overexpressed hMSCs and mMSCs were markedly higher than those in control cells, consistent with the observed increase in ALP activity (Figs. 1w and S2e). After 21 days, ARS staining showed enhanced mineral deposition in both SLC17A5(Slc17a5)-overexpressed hMSCs and mMSCs, with a significant increase in mineralized nodule formation (Fig. 1x and S2f). In hMSCs, the mRNA expression of osteogenic genes, including ALPL, COL1A1, SPP1, RUNX2, and SP7 (Fig. S1t–x), as well as the protein expression levels of COL-I, RUNX2, and OCN, were significantly upregulated (Fig. S1y). These findings indicate that Sialin promotes osteogenic differentiation in MSCs.
Mitochondrial localization of Sialin supports MSCs' osteogenesis by regulating mitochondrial functionMitochondria, as the energy center of all eukaryotic cells, play a crucial role in regulating the function of MSCs. Colocalization studies of Sialin and the mitochondrial marker, Mito Tracker, revealed that Sialin was localized to mitochondria in hMSCs (Fig. 2a), and that Sialin expression was significantly high in the mitochondrial fraction of SLC17A5-overexpressed hMSCs (Fig. 2b). To minimize crosstalk and perform an accurate colocalization analysis, we used two-color single-molecule localization microscopy to reveal the colocalization of Sialin with mitochondrial membrane TOMM20 in hMSCs (Fig. 2c).
Fig. 2
Sialin localizes in mitochondria and regulates osteogenic differentiation of MSCs via mitochondrial function. a IF staining images showing colocalization of Sialin (green) with mitochondria (MitoTracker, red) in hMSCs. Nuclei were counterstained with DAPI (blue). Scale bars, 10 μm. b Immunoblot analysis of Sialin expression in mitochondrial fractions from control and Sialin-overexpressing (HA-SLC17A5) hMSCs. c Single-molecule localization microscopy showing Sialin (red) colocalized with TOMM20 (green) in hMSCs. Scale bars, 1 μm. Quantification of JC-1 polymer/monomer ratio (d), ATP level (e), NAD+/NADH ratio (f), ROS production (g), and basal respiration (h) in mMSCs from 8-week-old WT and cKO male mice. IF staining images (i) and quantitation (j) of MitoTracker fluorescence in control and shSLC17A5 hMSCs. Scale bars, 25 μm. ATP level (k), NAD+/NADH ratio (l), ROS level (m), ATP production (n), maximal respiration (o), glycolytic capacity (p), and glycolytic reserve (q) in control and shSLC17A5 hMSCs. ALP staining and activity (r, s), and ARS staining with quantification of mineralized nodules (t, u) in shSLC17A5 hMSCs treated with CoQ10 and Sialin-overexpressing hMSCs treated with ROT. RT-qPCR analysis of BGLAP (v) and SPP1 (w) mRNA expression in shSLC17A5 hMSCs treated with CoQ10 and Sialin-overexpressing hMSCs treated with ROT. ATP production (x) and NAD+/NADH ratio (y) in shSLC17A5 hMSCs treated with CoQ10 and Sialin-overexpressing hMSCs treated with ROT. Data are presented as the mean ± SD, n = 3, except for j and s (n = 30). *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.000 1
Subsequently, Mito Tracker and JC-1 staining demonstrated that the fluorescence intensity (Fig. S3a) and JC-1 polymer/monomer ratio (Fig. 2d) were reduced in cKO mMSCs, suggesting that the mitochondrial membrane potential (MMP) decreased. The ATP level (Fig. 2e) and NAD+/NADH ratio (Fig. 2f) were significantly lower in the mitochondria of cKO mMSCs than those in the WT; however, the extent of ROS production increased (Fig. 2g). In addition, the oxygen consumption rate (OCR) of cKO mMSCs was assessed, and non-mitochondrial oxygen consumption and basal respiration were found to be significantly decreased (Figs. S3b and 2h).
Measurement of MMP using MitoTracker and JC-1 staining revealed a reduction in fluorescence intensity in SLC17A5 knockdown hMSCs compared to control cells (Fig. 2i, j), indicating impaired MMP. Consistently, intracellular ATP levels (Fig. 2k) and the NAD+/NADH ratio (Fig. 2l) were markedly decreased, whereas ROS levels (Fig. 2m) were significantly elevated in SLC17A5 knockdown hMSCs. Mitochondrial stress testing via OCR and extracellular acidification rate (ECAR) analysis revealed significantly reduced ATP production in SLC17A5 knockdown hMSCs compared with controls (Fig. 2n), suggesting diminished mitochondrial capacity to meet basal energy demands. Addition of the uncoupler carbonyl cyanide p-trifluoro-methoxyphenyl hydrazone (FCCP) further revealed that maximal respiration was significantly impaired in SLC17A5 knockdown hMSCs (Fig. 2o), indicating reduced bioenergetic responsiveness under stress conditions. Similarly, spare respiratory capacity was markedly decreased (Fig. S3c), further supporting a loss of metabolic flexibility. In mMSCs, basal respiration, ATP production, and maximal respiration were likewise significantly decreased following Slc17a5 knockdown (Fig. S3d–g). Interestingly, SLC17A5 knockdown hMSCs displayed an increase in both basal glycolytic capacity (Fig. 2p) and maximal glycolytic response (Fig. 2q), indicating a compensatory metabolic shift toward glycolysis.
Enhanced fluorescence intensity of MitoTracker staining (Fig. S3h, i) indicated that SLC17A5 overexpression increased MMP in hMSCs. Consistently, SLC17A5 overexpression significantly elevated intracellular ATP levels (Fig. S3j) and the NAD+/NADH ratio (Fig. S3k), while reducing ROS production (Fig. S3l). Basal respiration (Fig. S3m) and ATP production (Fig. S3n) were both significantly higher in SLC17A5-overexpressed hMSCs compared to control cells, suggesting an enhanced capacity for mitochondrial ATP synthesis to meet cellular energy demands. Upon FCCP treatment, maximal respiration was significantly increased in SLC17A5-overexpressed hMSCs (Fig. S3o), indicating improved bioenergetic responsiveness under stress conditions. Similar enhancements in basal respiration, ATP production, and maximal respiration were observed in Slc17a5-overexpressed mMSCs (Fig. S3p–s). Interestingly, glycolytic capacity (Fig. S3t) and glycolytic reserve capacity (Fig. S3u) were both reduced in SLC17A5-overexpressed hMSCs compared to controls, suggesting a metabolic shift favoring mitochondrial oxidative phosphorylation over glycolysis.
To determine whether Sialin influences the osteogenic differentiation potential of MSCs through regulation of mitochondrial homeostasis, we modulated mitochondrial electron transport chain (ETC) activity by pharmacological means: SLC17A5 knockdown hMSCs were treated with Coenzyme Q10 (CoQ10) to enhance ETC activity, while SLC17A5-overexpressed hMSCs were treated with rotenone (ROT) to inhibit it. After 7 days of differentiation, CoQ10 treatment enhanced ALP staining in SLC17A5 knockdown hMSCs, whereas ROT treatment led to a marked reduction in ALP staining intensity in SLC17A5-overexpressed hMSCs, with consistent changes observed in ALP activity assays (Fig. 2r, s). After 21 days, ARS staining showed increased mineralization and nodule formation in CoQ10-treated SLC17A5 knockdown hMSCs, whereas ROT-treated SLC17A5-overexpressed hMSCs displayed reduced mineral deposition (Fig. 2t, u). In parallel, qPCR analysis revealed significant upregulation of BGLAP and SPP1 in SLC17A5 knockdown hMSCs treated with CoQ10, and downregulation of these genes in SLC17A5-overexpressed hMSCs following ROT treatment (Fig. 2v, w). CoQ10 activation also restored ATP levels and the NAD+/NADH ratio in SLC17A5 knockdown hMSCs, while ROT treatment impaired both parameters in SLC17A5-overexpressed hMSCs (Fig. 2x, y). Collectively, these findings demonstrate that Sialin promotes osteogenic differentiation of MSCs by supporting mitochondrial function through regulation of ETC activity.
Mitochondrial-targeted Sialin overexpression enhances metabolic and functional homeostasis in MSCsTo specifically target Sialin to the mitochondria of hMSCs, we fused the mitochondrial targeting sequence derived from the lead peptide of cytochrome c oxidase subunit 8 (COX8) to the N-terminus of HA-tagged SLC17A5, generating a mitochondria-targeted Sialin construct (Fig. 3a, b). This mitochondrial-targeted overexpression significantly elevated Sialin protein levels in hMSCs (Figs. 3c, S3v). Following 7 days of osteogenic induction, hMSCs expressing mitochondria-targeted Sialin exhibited denser ALP staining than both control and conventional SLC17A5-overexpressed hMSCs (Fig. 3e). After 21 days, these cells also showed enhanced ARS staining and increased mineralized nodule formation (Fig. 3f). Consistently, the protein levels of osteogenic markers COL-I, RUNX2, Sialin, and OCN were upregulated in hMSCs with mitochondria-targeted SLC17A5 overexpression compared to both control and non-targeted SLC17A5-overexpressing cells (Fig. 3d).
Fig. 3
Mitochondria-targeted Sialin more effectively maintains MSCs homeostasis. a Schematic diagram of the construct for mitochondria-targeted overexpression of Sialin. b IF staining images showing colocalization of mitochondria-targeted Sialin (green) with mitochondria (MitoTracker, red) hMSCs. Nuclei were stained with DAPI (blue). Scale bars, 10 μm. c Immunoblot analysis of Sialin expression in mitochondrial fractions from control and mitochondria-targeted Sialin-overexpressing (HA-Mito-SLC17A5) hMSCs. d Immunoblot analysis of COL-I, RUNX2, Sialin, and OCN in control, Sialin-overexpressing (HA-SLC17A5), and mitochondria-targeted Sialin-overexpressing hMSCs. ALP staining and activity (e), and ARS staining with quantification of mineralized nodules (f) in control, Sialin-overexpressing, and mitochondria-targeted Sialin-overexpressing hMSCs. g IF staining images and quantitation of MitoTracker fluorescence intensity in control, Sialin-overexpressing, and mitochondria-targeted Sialin-overexpressing hMSCs. Scale bars, 25 μm. h OCR profiles of control, Sialin-overexpressing, and mitochondria-targeted Sialin-overexpressing hMSCs. Non-mitochondrial oxygen consumption (i), basal respiration (j), ATP production (k), and maximal respiration (l) in control, Sialin-overexpressing, and mitochondria-targeted Sialin-overexpressing hMSCs. ATP levels (m) and NAD+/NADH ratio (n) in control, Sialin-overexpressing, and mitochondria-targeted Sialin-overexpressing hMSCs. Data are presented as the mean ± SD, n = 3, except for g (n = 30). *P < 0.05; **P < 0.01; ***P < 0.001
MitoTracker fluorescence intensity was more markedly increased in hMSCs with mitochondria-targeted SLC17A5 overexpression than in those with non-targeted SLC17A5 overexpression (Fig. 3g), indicating a greater enhancement of MMP. We subsequently assessed the OCR in control, SLC17A5-overexpressed, and mitochondria-targeted SLC17A5-overexpressed hMSCs (Fig. 3h). Mitochondria-targeted SLC17A5 overexpression significantly increased basal respiration (Fig. 3j), non-mitochondrial oxygen consumption (Fig. 3i), and ATP production (Fig. 3k), even after ROT/antimycin A treatment. Following FCCP administration, maximum respiration was highest in hMSCs overexpressing mitochondria-targeted SLC17A5 (Fig. 3l), indicating an improved capacity to respond to elevated energy demand or metabolic stress. In addition, mitochondrial targeting led to a further increase in ATP levels (Fig. 3m) and the NAD+/NADH ratio (Fig. 3n) compared to both control and non-targeted SLC17A5-overexpressed hMSCs. Collectively, these results confirm that mitochondria-targeted SLC17A5 overexpression more effectively enhances mitochondrial homeostasis and promotes osteogenic differentiation of MSCs.
Sialin interacts with pSTAT3S727 and facilitates its mitochondrial translocationTo further explore the molecular mechanism by which Sialin regulates mitochondrial function, we extracted mitochondrial proteins from control and SLC17A5 knockdown hMSCs for phosphoproteomic analysis. PCA (Fig. S4a), combined with heatmap analysis, indicated significant differences in phosphorylation profiles between the two groups (Fig. S4b). A volcano plot of all the proteins in the differential phosphorylation analysis was generated, with red representing 116 significantly upregulated phosphoproteins and blue representing 226 significantly downregulated ones. Subsequently, the differentially phosphorylated proteins were subjected to gene ontology (GO) biological process (BP) enrichment (Fig. 4a), Reactome pathway (Fig. S4c), and WikiPathways analysis (Fig. S4d), which revealed that most of these proteins were involved in mitochondrial functions, various metabolic pathways, and signal transduction processes. These results support the critical role of Sialin in maintaining MSCs homeostasis by regulating mitochondrial function and cellular metabolism.
Fig. 4
Sialin interacts with pSTAT3S727 and promotes its mitochondrial translocation in MSCs. a Gene Ontology biological process (GO-BP) enrichment analysis of the differentially phosphorylated proteins. b Dual-view hierarchical graph learning model predicts the interaction between Sialin and candidate proteins. c Co-immunoprecipitation (Co-IP) showing the interaction between Sialin and pSTAT3S727 in hMSCs. d Surface plasmon resonance (SPR) analysis confirming the direct binding between Sialin and pSTAT3S727 in hMSCs (KD = 683 nmol/L). e IF staining images and quantitation of pSTAT3S727 colocalization with mitochondria by Pearson’s correlation coefficient in control, Sialin-overexpressing (HA-SLC17A5), and mitochondria-targeted Sialin-overexpressing (HA-Mito-SLC17A5) hMSCs. Scale bars, 10 μm. f A schematic model: Sialin binds pSTAT3S727, promotes its translocation to mitochondria, and thereby activates mitochondrial function in hMSCs. Data are presented as the mean ± SD, n = 3, except for f (n = 30). ****P < 0.000 1
Recently, a two-view layer hierarchical map learning model called HIGH-PPI was reported, which applies dual networks to represent protein structure and interaction interface structure, and achieves high accuracy and robustness in protein-protein interactions (PPIs) prediction.28 This model also enables identification of key binding sites and provides mechanistic interpretability. We applied the HIGH-PPI model to predict potential Sialin interactors among the differentially phosphorylated proteins and integrated these predictions with our phosphoproteomic data into a scatter plot (Fig. 4b), where the x-axis represents the HIGH-PPI–derived interaction score and the y-axis represents the change in phosphorylation level. This analysis highlighted pSTAT3S727 as the highest-scoring differentially phosphorylated protein associated with mitochondrial regulation, while other candidates exhibited low predicted binding capacity or lacked detectable interaction regions. Therefore, we chose pSTAT3S727 as the focus of subsequent studies. We confirmed that Sialin interacted with pSTAT3S727 in hMSCs, using classical co-immunoprecipitation (CoIP) (Fig. 4c). To clarify the binding strength of Sialin and pSTAT3S727, we used surface plasmon resonance (SPR) and found the binding constant (Ka) of 2.201E + 3, dissociation constant (Kd) of 1.505E-3, and affinity constant (KD) of 6.839E-7, which indicated strong binding (Fig. 4d). pSTAT3S727 affects mitochondrial function by enhancing the activity of the electron transport chain and increasing ATP production, and is critical for bone homeostasis by regulating osteogenesis.29,30,31 Furthermore, we found that the mitochondria-targeted SLC17A5 overexpression significantly promoted pSTAT3S727 expression in the mitochondria of hMSCs (Fig. 4e). In summary, these findings indicate that Sialin can interact with pSTAT3S727 and positively regulate the mitochondrial translocation of pSTAT3S727 in hMSCs to activate mitochondrial function (Fig. 4f).
Sialin-pSTAT3S727 axis controls MSCs osteogenesis and bone homeostasis via mitochondrial regulationSTAT3 has been considered as a potential target in several diseases, including cancer, immune disorders, and bone metabolism and homeostasis.32 Therefore, we wondered whether Sialin is a suitable pharmacological target for regulating bone homeostasis through pSTAT3. The classical STAT3 inhibitor, Stattic, inhibits the phosphorylation and activity of STAT3. To clarify the essential role of Sialin in regulating mitochondrial function, we inhibited STAT3 phosphorylation using Stattic and observed that MMP in hMSCs was significantly reduced and could no longer be enhanced by SLC17A5 overexpression (Fig. 5a). In line with this, cellular NAD+/NADH ratio and ATP levels were markedly decreased upon STAT3 inhibition, and SLC17A5 overexpression failed to restore them (Fig. S5a, b). Moreover, SLC17A5(Slc17a5) overexpression did not increase either basal or maximal respiration in Stattic-treated hMSCs and mMSCs, suggesting that pSTAT3S727 is required for Sialin-mediated enhancement of mitochondrial responsiveness under increased energy demand or stress (Figs. 5b, c and S5c, d).
Fig. 5
Inhibition of STAT3 phosphorylation abolishes the Sialin-induced enhancement of MSCs function and bone homeostasis. a IF staining images and quantitation of MitoTracker intensity in control and Sialin-overexpressing (HA-SLC17A5) hMSCs treated with the STAT3 phosphorylation inhibitor Stattic. Scale bars, 25 μm. Basal respiration (b) and maximal respiration (c) in control and Sialin-overexpressing hMSCs after Stattic treatment. d Representative micro-CT images of femurs. Upper scale bars, 1 mm. Lower scale bars, 200 μm. Quantification of BV/TV (e), BS/BV (f), Tb.N (g), and Tb.Sp (h) in femurs of control and Sialin-overexpressing mice treated with Stattic. H&E (i) and Masson’s trichrome staining (j) of femoral sections from control and Sialin-overexpressing mice treated with Stattic. Upper scale bars, 1 mm. Lower scale bars, 200 μm. IF staining images (k) and quantification (l) of Osx in femoral sections from control and Sialin-overexpressing mice treated with Stattic. Scale bars, 50 μm. Arrowheads indicate Osx-positive osteoblasts. Data are presented as the mean ± SD, n = 3, except for a (n = 30), d–l (n = 5). *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.000 1; n.s., no significance
Consistently, after 7 and 21 days of osteogenic induction, ALP staining intensity and activity, as well as ARS staining and mineralized nodule formation, were all reduced in Stattic-treated hMSCs and mMSCs compared to untreated controls (Fig. S5e–h). Importantly, SLC17A5(Slc17a5) overexpression did not rescue these osteogenic defects in the presence of Stattic (Fig. S5e–h). Nuclear counts using DAPI staining revealed comparable cell numbers between Stattic-treated and control groups, indicating that the observed reductions in ALP and mineralization were not due to cell loss (Fig. S5i). To further assess whether constitutive activation of STAT3 could compensate for Sialin deficiency, we expressed a phosphomimetic STAT3-S727D mutant in SLC17A5 knockdown hMSCs. STAT3-S727D partially restored ALP activity and osteogenic gene expression compared with SLC17A5 knockdown hMSCs transfected with empty vector, but the rescue remained incomplete relative to controls (Fig. S6a, b). To further explore the in vivo effects of pharmacological inhibition of STAT3 activity, 8-week-old mice were intraperitoneally injected with 10 mg/kg Stattic thrice per week for 4 weeks. Micro-CT analysis indicated that the trabecular bone was reduced by Stattic treatment, indicated by decreased BV/TV, BS/TV, and Tb.N and increased Tb.Sp, and overexpression of Sialin failed to restore bone homeostasis (Fig. 5d–h). In cortical bone, Sialin overexpression increased Ct.BV, Ct.Th, and Tt.Ar compared with vector controls, whereas Stattic treatment reduced these parameters (Fig. S6c–e). Sialin overexpression failed to rescue cortical deficits under Stattic treatment (Fig. S6c–e). The level of the bone resorption marker CTX-1 increased by Stattic treatment, and overexpression of Sialin failed to inhibit osteoclast activity and bone resorption status (Fig. S6f). The level of the bone formation marker PINP decreased by Stattic treatment, and overexpression of Sialin failed to enhanced osteoblast activity and bone formation status (Fig. S6g). In addition, hematoxylin and eosin (H&E) and Masson’s trichrome staining revealed structural disruption and decreased collagen content in the femur of the Stattic group, which also could not be restored by the overexpression of Sialin (Fig. 5i, j). IF staining showed that bones with overexpressed Sialin had many Osx+ osteoblasts (Fig. 5k, l). Osx+ osteoblasts were hardly observed in mouse bones after Stattic treatment, and overexpression of Sialin failed to increase the number (Fig. 5k, l). Quantification of TRAP staining in tibial sections showed that although Sialin inhibited basal osteoclast formation, it failed to suppress the Stattic-induced increase in osteoclast number (Fig. S6h). Taken together, these results suggest that Sialin regulates mitochondrial homeostasis via pSTAT3S727, thereby promoting osteogenic differentiation of MSCs and maintaining bone homeostasis.
Nitrate activates Sialin to restore MSCs' function and prevent age-related bone lossA positive feedback loop between nitrate and Sialin has been proposed to support nitrate-mediated regulation of nitric oxide (NO) homeostasis. In our study, nitrate treatment significantly upregulated Sialin expression in whole hMSCs (Fig. 6a) and specifically within the mitochondria (Fig. 6b), suggesting that nitrate may promote MSCs functional homeostasis via Sialin activation. Functionally, nitrate stimulation enhanced ALP staining intensity in hMSCs, with consistent increases observed in ALP activity (Fig. 6d), and markedly promoted mineralized nodule formation (Fig. 6e). The mRNA expression levels of osteogenic markers, including ALPL, BGLAP, SPP1, and RUNX2 (Fig. S7a–d), as well as the protein expression of COL-I, RUNX2, and OCN (Fig. 6c), were significantly upregulated following nitrate treatment. Moreover, nitrate treatment increased MMP, as evidenced by elevated MitoTracker fluorescence intensity (Fig. 6f), and improved mitochondrial redox status, as reflected by a higher NAD+/NADH ratio (Fig. 6g). OCR analysis further demonstrated that ATP production (Fig. 6h), maximal respiration (Fig. 6i), and spare respiratory capacity (Fig. 6j) were all significantly enhanced in nitrate-treated hMSCs.
Fig. 6
Nitrate-Sialin signaling is closely associated with senile osteoporosis in vivo. a Immunoblot analysis of Sialin expression in hMSCs treated with nitrate. b Immunoblot analysis of mitochondrial Sialin in hMSCs following nitrate treatment. c Immunoblot analysis of COL-I, RUNX2, and OCN in hMSCs treated with nitrate. ALP staining and activity (d), and ARS staining and mineralized nodule formation (e) in hMSCs treated with nitrate. f IF staining images and quantitation of MitoTracker fluorescence intensity in hMSCs treated with nitrate. Scale bars, 25 μm. Quantification of NAD+/NADH ratio (g), ATP production (h), maximal respiration (i), and spare respiratory capacity (j) in hMSCs following nitrate stimulation. Nitrate concentrations in saliva (k) and serum (l) of aged mice treated with nitrate. m Representative micro-CT images of femurs. Upper scale bars, 1 mm. Lower scale bars, 200 μm. Quantification of BV/TV (n) and Tb.N (o) in femurs from aged mice treated with nitrate. p Serum levels of PINP in aged mice following nitrate treatment. q, r, H&E (q) and Masson’s trichrome staining (r) in femoral sections from aged mice treated with nitrate. Upper scale bars, 1 mm. Lower scale bars, 200 μm. s IF staining images and quantification of Osx in femoral sections from aged mice treated with nitrate. Scale bars: 50 μm. Arrowheads indicate Osx-positive osteoblasts. Data are presented as the mean ± SD, n = 3, except for f (n = 30), except for k, l, n–p, s (n = 5). *P < 0.05; **P < 0.01; ***P < 0.001
To further explore the preventive effect of nitrate–Sialin on age-induced osteoporosis in vivo, we successfully established a model of senile osteoporosis in 18-month-old mice and administered 4 mmol/L nitrate in water for 6 months (Fig. S7e). Nitrate concentrations in the saliva and blood of aged mice significantly increased by nitrate administration (Fig. 6k, l). Micro-CT scanning of the femur microstructure (Fig. 6m) revealed that the BV/TV and Tb.N were elevated while Tb.Sp was decreased after nitrate administration (Fig. 6n, o and Fig. S7f), indicating that nitrate significantly elevated bone volume in aged mice. In aged mice, nitrate administration increased cortical Ct.BV, Ct.Th, and Tt.Ar compared with aged mice, consistent with the trabecular improvements (Fig. S7i–k). The CTX-1 level was significantly reduced in mouse blood after nitrate administration, indicating that nitrate inhibited osteoclast activity and bone resorption status in aged mice (Fig. S7g). The P1NP level was significantly increased in mouse blood after nitrate administration, indicating that nitrate enhanced osteoblast activity and bone formation status in aged mice (Fig. 6p). In addition, H&E and Masson’s trichrome staining revealed improved bone architecture and increased collagen content in aged mice after nitrate administration (Fig. 6q, r). IF staining showed a significant increase in the number of Osx+ osteoblasts in the bones of senescent mice (Fig. 6s). Quantitative analysis of TRAP staining in tibial sections showed that nitrate administration significantly reduced osteoclast numbers in aged mice (Fig. S7h). After 21 days of osteogenic induction, ARS staining intensity and mineralized nodule formation were significantly increased in mMSCs isolated from aged mice following nitrate administration (Fig. S7l). In parallel, the mRNA expression levels of Alpl, Spp1, an
Comments (0)