
Remember me
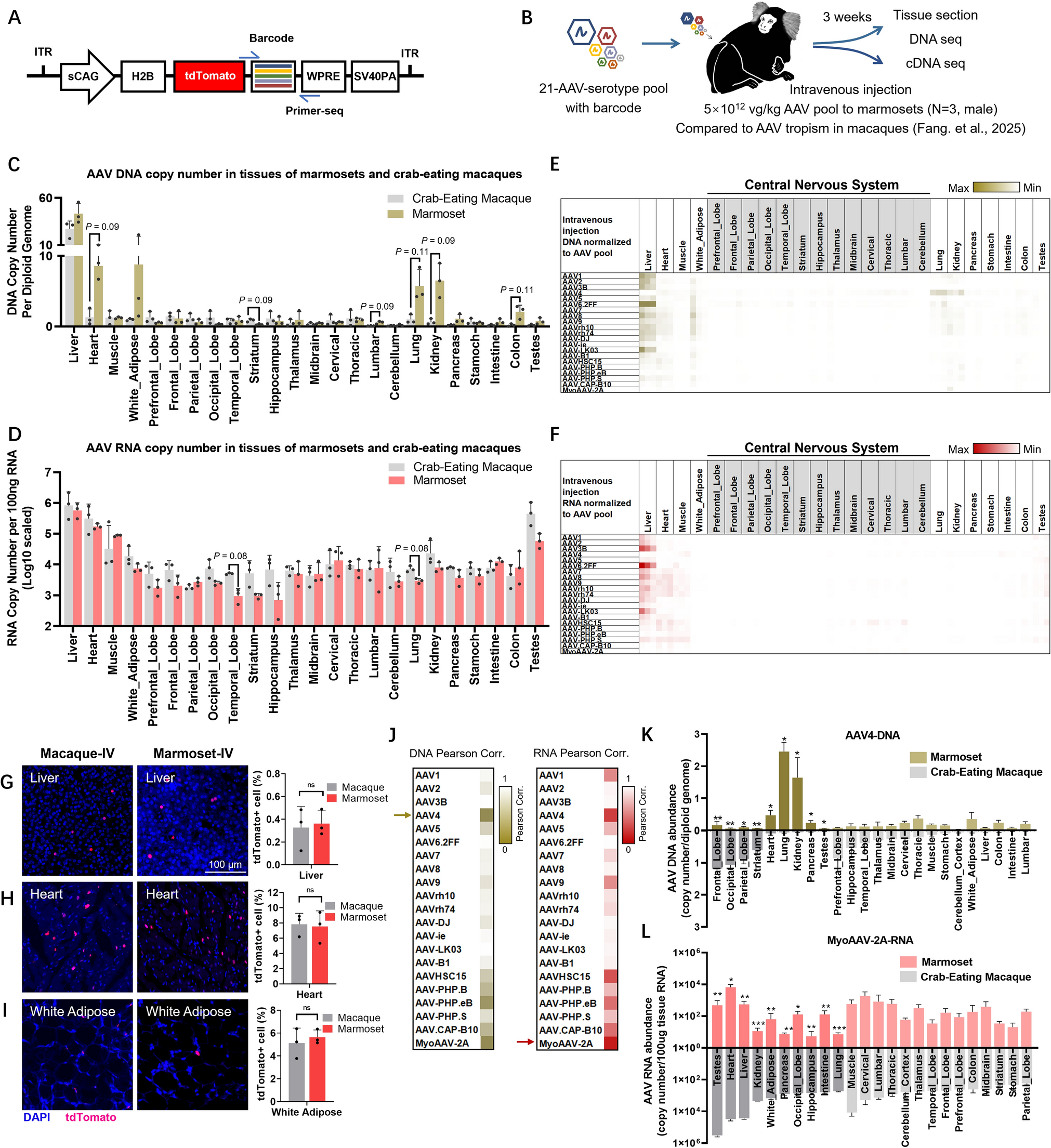

To test whether previously learned fear can still alter visual processing in a safe context, we first established a fear conditioning task (Fig. 1A) in a threatening context in Experiment 1, then switched to a safe context (Fig. 1B)—rendering learned threat cues invisible and incorporating these subliminal cues into the main task (Fig. 1C).
In the fear conditioning task of Experiment 1 (threat context, Fig.1A), luminance gratings with either clockwise or counterclockwise orientations were used. For each participant, one orientation (CS+, 100% contingency) was repeatedly paired with electric aversive shocks (rated 7–8 on a scale of 0 to 10, where 0 indicated no pain at all and 10 indicated extreme pain). The other grating, with an orthogonal orientation, was never paired with shocks (CS−).
Subsequently, the main task switched to a safety context (Fig. 1B). To ensure a safe context, participants were explicitly informed that no further electric shocks would occur, and they practiced the main task beforehand to confirm this safety. Furthermore, we used a different stimulus set in the main task. That is, black-and-white luminance gratings were replaced with red-and-green chromatic gratings. These chromatic gratings were rapidly alternated with their anti-phase counterparts to achieve chromatic flicker fusion. Under these conditions, gratings of different orientations were consciously perceived as identical yellow disks (Fig. 1C)—a significant difference from the tilted black-and-white gratings perceived during fear conditioning.
To verify that orientation information was processed unconsciously during the main task and to direct attention to the CFF stimuli, we presented CFF stimuli in the central visual field (where attention is most concentrated). Participants were required to perform a 2-alternative forced choice (2-AFC) task (Fig.1C), indicating which flicker was in clockwise or anti-clockwise orientation. Task performance confirmed that participants could not discriminate grating orientation during the main task: after excluding one participant whose 2-AFC discrimination accuracy (ACC) reliably exceeded the 50% chance level (77.5% and 70% in Sessions 1 and 2, respectively), the remaining participants performed at chance (Session 1: t(14) = −0.09, P = 0.930, accuracy range: 0.375–0.650; Session 2: t(14) = −0.95, P = 0.357, accuracy range: 0.350–0.550; Fig. 1A). Only data from these remaining 15 participants were included in subsequent analyses of Experiment 1.
We hypothesized that previously established fear associations would generalize to the safe context and subsequently alter stimulus-evoked visual responses unconsciously. To better evaluate the temporal profile of such visually tagged unconscious fear responses in the safe context, CFF stimuli were presented for up to 2 seconds. This allowed continuous monitoring of responses at stimulus onset, offset, and during stimulus maintenance.
We first sought physiological evidence of unconscious fear in the safe context. Previous studies have shown that unconscious fear can be indexed by eye-related measures, such as increased pupil diameter and electromyography (EMG) signals recorded from electrodes above the left eyebrow [21, 32]. This inspired us to examine the EEG signals simultaneously recorded by bipolar electrodes for vertical electrooculography (VEOG), which were intentionally placed above (VEOU) and below (VEOL) the left eye (Fig. 1E left panel; see also Methods). We hypothesized that if unconscious fear occurred, fear-related information would induce a differential VEOG signal between unconscious CS+ and CS−. To control for volume conduction, we further subtracted the mean signal of three frontal electrodes surrounding the left eye (i.e., FPZ, FP1, F7, which showed distinct raw signal patterns) from the VEOG signal. As expected, we observed a significant difference between CS+ and CS− in the resulting signal (353–614 ms, summed T = 69.57, corrected P = 0.01; Fig. 1E right panel). Since all other aspects of the CSs were identical except for their threat-related associations, this unique electrooculogram response provides additional physiological evidence for the occurrence of unconscious fear.
Our primary interest was to determine whether unconscious fear alters visual processing in the safe context. We hypothesized that unconscious fear facilitates visual processing, potentially leading to differential event-related potentials (ERPs) between invisible CS+ and CS− flickers—particularly at occipital electrodes. To test this, we focused our analysis on an occipital region of interest (occipital ROI, Fig. 1F, middle upper panel) including six electrodes O1, Oz, O2, PO3, POz, and PO4. Using the mean ERP amplitude averaged across these six electrodes as the dependent variable and applying a cluster-based permutation test for multiple comparison correction, we identified three temporal clusters with significant differential ERPs (Fig. 1F left panel): 383–865 ms (summed T = 159.4, corrected P = 0.002), 1196–1547 ms (summed T = 118.1, corrected P = 0.012), and 1717–1888 ms (summed T = 46.9, corrected P = 0.040). To further characterize these findings, we assessed the spatial distribution of the mean differential ERP averaged across each cluster, confirming that the aforementioned unconscious fear responses were predominantly localized to occipitoparietal regions (Fig. 1F right panel). In contrast, the same analysis revealed no significant fear responses in frontal, central, or parietal ROIs (Fig. S1, note that the parietal ROI exhibited a similar response pattern to the occipital ROI, but failed to survive multiple comparison correction).
To investigate which neural oscillations mediate the communication of unconscious fear signals, we performed an inter-trial coherence (ITC) analysis—restricting it to slow neural oscillations (3–30 Hz), as these are better suited for inter-regional long-distance communication than fast oscillations [33,34,35]. Consistent with the ERP findings, we first conducted this analysis in the occipital ROI. Temporally, three significant ITC clusters (Fig. 1G) were identified at 98–487 ms (cluster-based permutation test, summed T = 370.1, corrected P = 0.004), 1421–1621 ms (summed T = 248.7, corrected P = 0.030), and 1727–1982 ms (summed T = 192.4, marginally significant at corrected P = 0.058). In sharp contrast, no significant ITC effects were observed in frontal, central, or parietal ROIs (Fig. S2). To identify the frequency band most affected by unconscious fear, we conducted a paired-sample t-test comparing CS+ and CS− in the occipital ROI, using the mean ITC value averaged across 100–2000 ms post-stimulus onset as the dependent variable. Results showed significantly stronger ITC for CS+ than CS− in the high theta band (6–8 Hz), peaking at 7 Hz (t(14) = 4.62, uncorrected P < 0.001, FDR-corrected P < 0.05; Fig. 1H).
Although both ERP and ITC effects reflect unconscious fear processing, they exhibit distinct temporal and spatial patterns. Temporally, ITC effects are mainly confined to stimulus onset and offset, with only transient overlap with differential ERP responses (Fig. 1I). Spatially, ITC effects are concentrated at midline occipital electrodes (POz and Oz; Fig. 1G, three nested topographies), showing a more focused distribution than ERP effects. These temporal and spatial differences provide preliminary evidence that theta-band ITC effects and differential ERP effects represent two distinct neural processes underlying unconscious fear processing (see Discussion).
Collectively, the results of Experiment 1 demonstrate that previously learned associative fear can unconsciously enhance visual responses (i.e., distort vision) even in a safe context after the imminent threat has been removed. This distorted visual processing is represented by two distinct neural correlates in human occipital regions: visually evoked ERPs and theta coherence (ITC) peaking at 7 Hz.
Unconscious Fear Requires Attention to Distort Visual ProcessingFollowing the findings of Experiment 1, we next asked: Is the visually-tagged unconscious fear response in the safe context purely stimulus-driven—distorting primary visual processing without requiring higher-order cognitive factors? Among such cognitive factors, top-down attention is characterized by its susceptibility to cognitive control and is a core target of current mainstream therapies for anxiety disorders [12,13,14]. Previous literature also identifies top-down attention as a fundamental factor modulating a wide range of unconscious processes related to language, vision, and emotion (for a review, see [36]). Clarifying whether attention is required for unconscious fear to distort vision in the safe context is critical for evaluating the efficacy of attention-based therapies in regulating such unconscious fear responses.
To address this question, Experiment 2 focused on top-down attention and explored its impact on the unconscious fear responses observed in Experiment 1. This research objective required unconscious fear cues to be displayed in an attended state and an unattended state, respectively, with comparisons between the two conditions to reveal the regulatory effect of attention.
To create these two attention conditions, we moved the subliminal fear cues originally presented in the central visual field to the peripheral visual field (Fig. 2A). Similar to Experiment 1, each trial still presented one CS+ and one CS- in random order, but with a key difference: both the CS+ and CS- were composed of a pair of two symmetric chromatic gratings of the same orientation (Fig. 2A). This symmetric presentation was used during both the fear conditioning task when grating orientations were explicitly visible (Fig. S3) and the main task when chromatic gratings were rendered invisible via CFF. This symmetrical design offers two critical advantages: first, compared to a single stimulus, symmetric presentation enhances visual input intensity, making participants’ neural responses stronger and easier to detect attention-induced modulations; second, this symmetrical presentation balances visual salience on both sides, eliminating the bias toward one side that may occur with unilateral stimuli. This helped reduce visual distraction and related visual saccades, thus maintaining stable central fixation and ensuring chromatic flicker fusion.
The specific task under the attended condition was a duration comparison task: CS+ pairs and CS- pairs were presented in random order, with each grating pair having one of three possible durations (800, 1000 and 1200 ms, Fig. 2B). The durations of the two grating pairs in each trial were always different, and participants were required to focus on distinguishing which pair had a longer duration. The experimental results showed that participants’ accuracy in this task was significantly higher than the chance level (accuracy: 0.73 ± 0.11, t(22) = 10.41, P < 0.01), demonstrating that they effectively engaged in the task and successfully focused their attention on the subliminal fear cues in the peripheral visual field.
Under the unattended condition, participants performed a color detection task: they were instructed to ignore the CS+ and CS- pairs in the peripheral visual field and solely monitor the frequency of color changes of the central fixation point. The frequency of color changes per trial was randomized (0, 1, or 2, Fig. 2B), and participants were required to provide accurate counts. High task accuracy (accuracy: 0.94 ± 0.06, t(22) = 52.26, P < 0.001) confirmed that participants maintained robust central attention, effectively leaving peripheral CFF stimuli in an unattended state.
Prior studies [37,38,39] have demonstrated that the human brain is capable of unconsciously processing orientation information and inducing the orientation adaptation aftereffect—namely, the tilt-after effect (TAE). Critically, the TAE elicited by unconscious orientation information could be impervious to attentional modulation [37]: it occurs consistently under both attended and unattended conditions, with no significant difference in effect magnitude observed between the two attentional states (but also note that such TAE effect induced by invisible orientation processing could increase with attention under certain carefully designed conditions [38]). From this perspective, our study adopted an experimental design in which orientation information was fused to invisible stimuli and tested under both attended and unattended conditions. Therefore, any observed differences between these two attentional states can only reflect the processing of fear associated with invisible orientation cues, rather than loss of the brain’s inherent capacity to decode orientation information itself under unattended conditions.
Based on the two tasks described above, another key methodological advancement of Experiment 2 over Experiment 1 lies in the elimination of task-relevance confounds (also see Discussion). In Experiment 1, participants completed an orientation discrimination task, which was inherently task-relevant to the core associative feature (orientation) of fear conditioning—introducing potential interference despite the subliminal nature of the orientation cues. In contrast, both the duration discrimination task (attended condition) and central fixation color change detection task (unattended condition) in Experiment 2 were orthogonal to the orientation of the CS+ and CS- pairs. This design ensures that observed differences between the two attention conditions cannot be attributed to task-relevance effects, but rather reflect the intrinsic link between top-down attention and unconscious fear responses in safe contexts.
We first compared the intensity of stimulus-evoked visual responses under attended and unattended conditions. To quantify this intensity, we calculated the mean peak-to-peak amplitude of the N2 and P3 components of stimulus-evoked vERPs (average of CS+ and CS−, Fig. 2C left panel) calculated using bilateral electrodes in lateral occipitoparietal regions (Fig. 2C right panel). For early vERPs (~400ms post-stimulus onset), the unattended condition and the attended condition shared comparable peak-to-peak amplitude (t(22)= 1.21, P =0.240, Fig. 2C middle panel)—ruling out the possibility of significant suppression of early visual activity due to lack of attention in the unattended condition. As shown, the distribution of peak-to-peak amplitude across all scalp electrodes is consistent with the retinotopic activation pattern, wherein bilateral peripheral stimuli elicit activation in the lateral occipital electrodes. Accordingly, this symmetric peripheral ROI (Fig. 2C right panel) was adopted for all subsequent analyses in Experiment 2.
ERP analysis in the attended condition (Fig. 2E) revealed significant differential vERPs (CS+ vs. CS−) in two clusters: 196–452 ms (summed T = 159.5, corrected P = 0.042) and 532–1116 ms (summed T = 122.2, corrected P = 0.002) post-stimulus onset. At a cluster threshold of P < 0.05, CS+ induced significantly greater ERP responses than CS−. Additionally, 88–136 ms post-CS onset, CS+ induced significantly higher average ERP amplitude than CS−, though this effect did not survive multiple comparison correction (summed T = 31.0, uncorrected P < 0.05, corrected P = 0.392). These results demonstrate prolonged differential ERP representation during stimulus presentation, replicating unconscious fear effects under attended conditions.
ERP analysis in the unattended condition (Fig. 2D), however, revealed no significant vERP-based unconscious fear response between unconscious CS+ and CS- in each of the three clusters previously identified in the attended condition. Furthermore, the contrast between attended vs. unattended conditions using one-tailed paired t-test revealed the amplitude of this unconscious fear response decreased if attention was directed away (88–136 ms: t(22)=-1.54, P =0.068; 196–452 ms: t(22)=−1.81, P =0.042; 532–1116 ms: t(22)=-1.53, P =0.070). Results above thus provide compelling evidence showing that unconscious fear responses in safe contexts are present in the attended condition only but completely absent in the unattended condition.
In sharp contrast, ITC analysis revealed no significant ITC differences between unconscious CS+ vs. CS− under both unattended (Fig. 2F) and attended (Fig.2G) conditions in Experiment 2 (cluster-based permutation test, all corrected P > 0.05)—a notable departure from Experiment 1.
Collectively, the above findings provide compelling evidence that top-down attention can modulate unconscious fear responses in a safe context, thus extending attention’s regulatory effects on conscious stimulus processing (i.e., facilitating the processing of some stimuli while inhibiting that of others) to a broader spectrum of unconscious processes represented by unconscious fear (Fig. 2H).
Attention-Dependent Broad-Alpha Activity Bidirectionally Regulates Unconscious FearWhy was the vERP-based unconscious fear response present in the attended condition but completely absent in the unattended condition According to theoretical accounts [26], alpha oscillations can bidirectionally modulate unconscious processes prior to the emergence of perceptual awareness, serving two opposing functions to enhance signal-to-noise ratio (SNR). On one hand, alpha oscillations can suppress certain unconscious processes by eliminating them as noise, with higher alpha power enabling more effective noise reduction. On the other hand, they can facilitate specific unconscious processes by amplifying the signal information that is poised for expression, such that greater alpha power correlates with enhanced signal strength. It is proposed that the brain employs this alpha-band bidirectional modulation to amplify target signals and suppress irrelevant noise, thereby selecting emerging signals that ultimately gain access to conscious experience. However, empirical evidence supporting this bidirectional regulatory role of alpha oscillations in unconscious processes remains scarce.
In the present study, the pattern of unconscious fear effects across the two attentional conditions aligns precisely with the framework of signal enhancement under attention and noise suppression in the absence of attention. Guided by this insight, we further investigated the relationship between alpha oscillations and unconscious fear responses in each attentional condition.
We first tested the possibility that top-down attention suppresses unconscious fear via alpha oscillations under the unattended condition (Fig. 3A, Hypothesis 1). As alpha oscillations are thought to mediate suppression of the peripheral visual field [40,41,42], we hypothesized that post-stimulus alpha power within the lateral visual ROI might be elevated in the unattended condition relative to the attended condition. To address this hypothesis, we computed neural oscillation power across a broad frequency range and compared the unattended and attended conditions.
Fig. 3
Hypotheses and supporting evidence illustrating how top-down attention recruits alpha oscillations to bidirectionally modulate unconscious fear processing. A Hypothesis 1 posits that robust post-stimulus alpha-band oscillations are induced under the unattended condition, which suppresses unconscious fear processing by treating it as neural “noise”. B Empirical evidence for Hypothesis 1. White-outlined regions indicate statistically significant increases in broad-alpha power under the unattended versus attended condition. The red square marks the resulting region of interest (ROI) in the broad alpha band, which covers the classical alpha band (6–13 HZ) with broader band width, and was subsequently extracted and used for downstream statistical analyses. C Hypothesis 2 proposes that broad alpha oscillations facilitate the encoding and propagation of survived ‘signals’ under the attended condition, thereby correspondingly amplifying unconscious fear responses in this context. Pearson correlation analyses of data from Experiment 2 corroborated this hypothesis, revealing a significant positive correlation between post-stimulus broad alpha power and the amplitude of unconscious fear responses across all included participants. D Hypothesis 2 received additional empirical support from the data of Experiment 1. For Panels C and D, black dots denote individual participant data included in the correlation analyses, while gray dots represent outliers (normalized Z > 3 or < −3) that were excluded from the correlation analyses to ensure statistical validity. † P < 0.1; * P < 0.05; ** P < 0.01.
Consistent with this prediction, we observed stronger broad-alpha activity (4–20 Hz, Fig. 3B, red square) following CS onset in the unattended condition. This increase in alpha power emerged at around 200 ms post-stimulus and persisted until stimulus offset—a time window that overlaps with the period of significant unconscious fear responses in the attended condition (Fig. 2E). This temporal correspondence suggests that the heightened and sustained alpha activity suppressed unconscious fear responses. Framed within the signal-noise framework, the unconscious fear response in the attended condition (construed as a signal) was eliminated as noise by alpha oscillations in the unattended condition (i.e., a noise reduction process), thereby preventing the manifestation of differential vERP responses. Importantly, paired t-tests revealed no significant difference in broad-alpha oscillation power between CS+ and CS− stimuli (paired t-test, 196-452ms: t(22)=0.72, P =0.480; 532-1116ms: t(22)=1.28, P =0.214). Collectively, these findings demonstrate that broad-alpha oscillations do not directly encode unconscious fear (i.e., they do not differentiate between CS+ and CS−) but instead abolish unconscious fear responses in the unattended condition.
We next examined whether top-down attention facilitates signal expression via alpha oscillations in the attended condition (Fig. 3C, Hypothesis 2). The robust unconscious fear response observed in the attended condition indicates that this process was prioritized as a signal rather than suppressed as noise. Here, we specifically tested whether the strength of this signal would correlate positively with broad-alpha power. We predicted that if unconscious fear were treated as a prioritized signal and amplified by alpha-band activity, a positive correlation between these two measures would be observed.
To evaluate this prediction, we computed differential ERP amplitudes (CS+ minus CS−) to index unconscious fear and averaged post-stimulus alpha-band power (across CS+ and CS−) to quantify alpha activity for each participant. A Pearson correlation analysis was then conducted to assess the relationship between these two metrics. As shown in Fig. 3C, results revealed a significant positive correlation between differential ERP amplitudes and mean broad-alpha power during the two temporal clusters where unconscious fear responses were most prominent (Cluster 1: 196–452 ms, r = 0.45, P = 0.040; Cluster 2: 532–1116 ms, r = 0.43, P = 0.048). It is critical to note, however, that this positive correlation could potentially be confounded by a scaling effect—a phenomenon in which the magnitude of a condition-dependent effect varies with the baseline activity level. To rule out this confounding factor, we adopted the mean peak-to-peak amplitude of the N2 and P3 components of stimulus-evoked vERPs (Fig. 2C left panel) as a baseline index to quantify baseline activity; notably, no significant correlation (Fig. S4) was detected between this alternative baseline index and the unconscious fear effect of interest. This result confirms that the observed positive correlation between the unconscious fear and mean alpha oscillatory power is not attributable to a scaling effect, but rather represents a genuine association between the two variables.
Collectively, above results support the hypothesis that unconscious fear is prioritized as a signal and is thus protected or facilitated by alpha-band activity in the attended condition, providing direct empirical evidence that attention can recruit alpha oscillations to enhance unconscious processes such as unconscious fear.
To validate the reliability of this novel finding, we cross-validated the results across Experiment 1 and Experiment 2, testing whether the alpha-mediated facilitation of unconscious fear responses could generalize to the experimental paradigm of Experiment 1. Unlike Experiment 2 (which used bilateral peripheral CFF flickers and covert attention with a peripheral discrimination task), Experiment 1 presented CFF flickers in the central visual field, required overt attention to the flickers, and employed an orientation discrimination task (with orientation stimuli rendered invisible via perfect fusion). We replicated the same analytical pipeline to quantify unconscious fear responses and post-stimulus alpha-band activity, and observed a consistent positive correlation (Fig. 3D) between these two measures across all three temporal clusters previously reported in Fig. 1F (Cluster 1: 383–865 ms, r = 0.48, P = 0.072, marginally significant; Cluster 2: 1196–1547 ms, r = 0.59, P = 0.027; Cluster 3: 1717–1888 ms, r = 0.74, P = 0.002). Despite differences in stimulus presentation, attentional demands, and task design across the two experiments, this cross-experimental replication provides compelling evidence that attention reliably recruits broad-alpha oscillations to facilitate unconscious fear processing if these subliminal cues were in attended state.
Comments (0)