
Remember me
To comprehensively elucidate the molecular and functional consequences of STAT5A disruption in chronic myeloid leukemia (CML), we integrated bioinformatic, genetic, and phenotypic analyses across parental and tyrosine kinase inhibitor (TKI)-resistant K562 models. The results are presented in a multi-layered framework encompassing (i) transcriptomic profiling of the STAT5 signaling network using public CML datasets, (ii) functional validation of CRISPR/Cas9-mediated STAT5A knockout, and (iii) downstream phenotypic characterization including cell viability, apoptosis, and cell-cycle modulation. This integrative approach enables a mechanistic dissection of STAT5A-dependent resistance pathways and clarifies how targeted gene editing reprograms survival and apoptotic circuits in TKI-refractory CML cells.
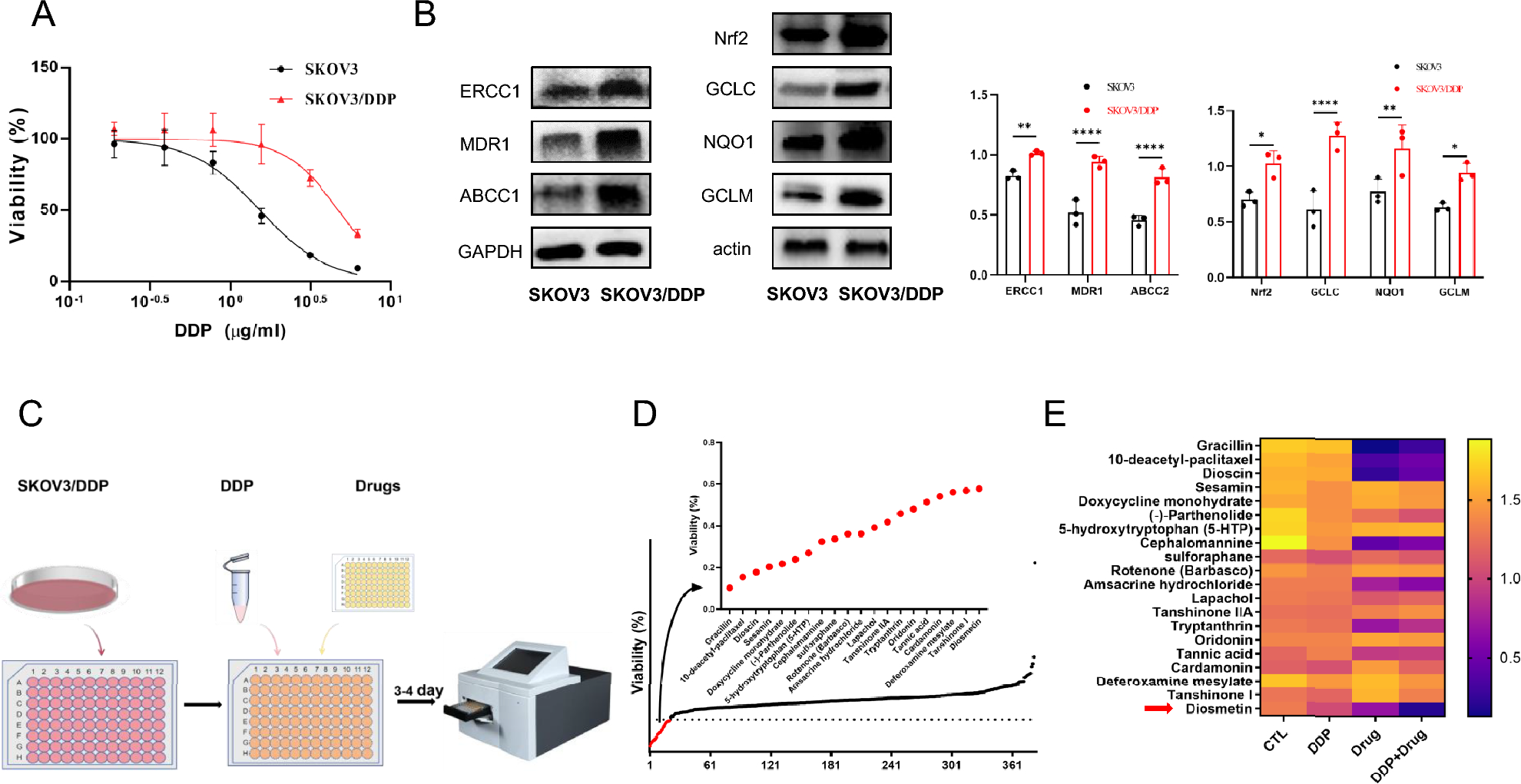
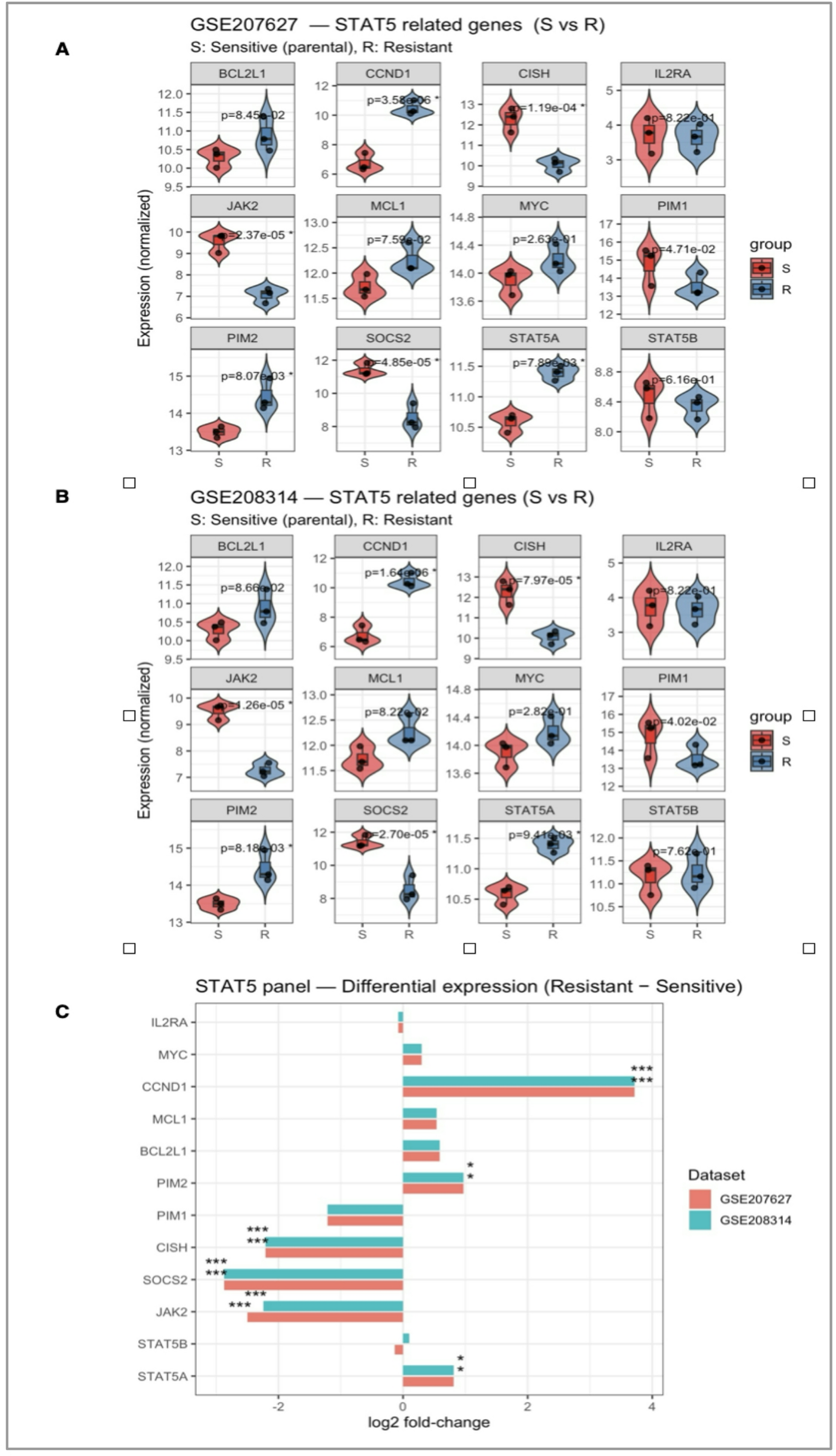
Transcriptomic remodeling of the STAT5 network underlies tyrosine kinase inhibitor resistance in chronic myeloid leukemiaTo explore the molecular basis of tyrosine kinase inhibitor (TKI) resistance, we first examined transcriptomic changes across two independent datasets (GSE207627 and GSE208314). Across two independent datasets (GSE207627 and GSE208314), consistent transcriptional alterations were detected within the STAT5 signaling network (Fig. 1). Both STAT5A and STAT5B, together with the upstream kinase JAK2, were markedly downregulated in resistant clones, suggesting attenuation of canonical STAT5 activity upon chronic TKI exposure. In contrast, MCL1, CCND1, and BCL2L1—genes associated with survival and proliferation—were upregulated, implying compensatory activation of alternative pro-survival pathways. Notably, negative feedback regulators SOCS2 and CISH exhibited concordant suppression, indicating a breakdown in the autoregulatory control of STAT5 signaling.
Collectively, these patterns suggest a transcriptional rewiring from a STAT5A-driven regulatory circuit toward alternative survival mechanisms, providing a mechanistic basis for resistance acquisition. This observation guided subsequent analyses focusing on the functional and prognostic relevance of STAT5A in TKI-resistant CML.
Fig. 1 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Transcriptomic comparison of STAT5-associated signaling in TKI-sensitive and -resistant CML cells. Differential expression analysis of STAT5-related genes (STAT5A, STAT5B, JAK2, SOCS2, CISH, MCL1, CCND1, BCL2L1, IL2RA, PIM1, PIM2, MYC) across TKI-sensitive (S) and -resistant (R) CML datasets. A Expression patterns in GSE207627 (S vs. R). B Validation of the same gene panel in GSE208314. C Comparative log2 fold-change values of each gene across both datasets (resistant vs. sensitive). A shared color legend is applied for all panels (red = sensitive/parental; blue = resistant). Significance levels: p < 0.05 (*), p < 0.01 (**), p < 0.001 (***)
Baseline viability and cytotoxic response profiles of parental and TKI-resistant K562 cellsThe viability of cultured K562, K562/1.2 µM imatinib-resistant, and K562/3 nM ponatinib-resistant cells was assessed by trypan blue staining. The analysis showed that K562 cells had a viability of 95%, imatinib-resistant cells 92%, and ponatinib-resistant cells 95%. These results indicate that all cell groups had sufficient viability for subsequent experimental studies. To investigate the cytotoxic effects and the response of the resistant cell lines to the drug, an XTT assay was performed. K562/1.2 µM imatinib-resistant cells were treated with 0.5, 1, 1.2, and 2 µM imatinib, while K562/3 nM ponatinib-resistant cells were treated with 1, 3, and 6 nM ponatinib. The data obtained were analyzed using Calcusyn software, and the IC₅₀ values were calculated. After 48 h, the IC₅₀ value of the imatinib-resistant cells was 2.4 µM, while the ponatinib-resistant cells had an IC₅₀ value of 6 nM (Fig. 2A and B).In addition, drug response curves generated for parental and resistant cell lines showed marked differences in sensitivity to TKI treatment (Fig. 2C–F). Parental K562 cells displayed a pronounced dose- and time-dependent reduction in viability after exposure to both imatinib and ponatinib (Fig. 2C and E). In contrast, K562/1.2 µM imatinib-resistant cells showed reduced sensitivity to imatinib, as indicated by a rightward shift in the dose–response curve and increased IC₅₀ values compared to parental cells (Fig. 2D). Similarly, K562/3 nM ponatinib-resistant cells exhibited diminished responsiveness to ponatinib, requiring higher drug concentrations to achieve comparable cytotoxic effects, with an IC₅₀ value exceeding the ponatinib concentration used for resistance selection (Fig. 2F).
Fig. 2 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Cytotoxicity profiles of TKI-resistant chronic myeloid leukemia models. A, B XTT viability assay results for K562/1.2 µM imatinib-resistant and K562/3 nM ponatinib-resistant cells following 48 h drug exposure. Dose-dependent reductions in cell viability were used to determine IC₅₀ values. Data represent mean ± SD from three independent experiments; IC₅₀ values were calculated using CalcuSyn 2.0 software. C–F Dose–response curves of parental and resistant K562 cell lines to TKIs. Parental K562 cells exhibited a marked dose- and time-dependent decrease in viability in response to imatinib (C) and ponatinib (E). K562/1.2 µM imatinib-resistant cells (D) displayed reduced sensitivity to imatinib, as indicated by a rightward shift in the dose–response curve. K562/3 nM ponatinib-resistant cells (F) showed diminished responsiveness to ponatinib, with an IC₅₀ exceeding the concentration used for resistance selection. All data are presented as mean ± SD and were analyzed using GraphPad Prism (n = 3)
Lipofectamine CRISPRMAX-Mediated Delivery Enables Efficient CRISPR/Cas9 Genome Editing of STAT5A in K562 CellsK562 cells (2.5 × 105) were seeded in 6-well plates and transfected with guide RNAs targeting STAT5A, CDK4, and BRD4 along with Cas9 protein using Lipofectamine CRISPRMAX. After 48 h, cells were harvested, and genome editing efficiency was assessed using the GeneArt™ Genomic Cleavage Detection Kit. Gel electrophoresis analysis showed successful cleavage at the target loci. Targeting exon 2 of STAT5A (target sequence: AAGTAGTGGCCGGACCTCGAT, PAM: GGG) resulted in distinct bands at 271 bp and 220 bp. In the positive control (CDK4), cleavage products were observed at 557 bp, 342 bp, and 235 bp. No cleavage or additional bands were detected in the negative control group (non-targeted gRNA) Supplementary Figs. 1 and 2. Indel analysis revealed a cleavage efficiency of 18.8% in STAT5A knockout cells and 19.2% in CDK4 knockout cells (Fig. 3B). No significant cleavage was observed for BRD4. These results demonstrate that the CRISPR-Cas9 system can be efficiently introduced into K562 cells with Lipofectamine CRISPRMAX, enabling precise gene silencing at the desired sites (Fig. 3).
Fig. 3 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.CRISPR/Cas9 targeting of the STAT5A gene and indel efficiency analysis. A CRISPR/Cas9 targeting site within exon 2 of the STAT5A gene. The red sequence indicates the gRNA target region, and the triangle denotes the PAM site. B Indel frequencies (%) were quantified 48 h post-transfection. Non-targeting gRNA (NT gRNA) was used as a negative control, and CDK4 gRNA served as a positive control for editing efficiency. Data represent mean ± SD from three independent experiments (NT gRNA: 0.38 ± 0.03; STAT5A gRNA: 18.80 ± 0.41; CDK4 gRNA: 19.23 ± 0.55). Statistical significance was determined by one-way ANOVA followed by Dunnett’s multiple comparisons test versus the non-targeting gRNA control (****p < 0.0001). Full, uncropped gel and blot images corresponding to previous panels (C–D) are provided in Supplementary Figures S1–S2
To confirm that CRISPR/Cas9-mediated genome editing effectively depleted STAT5A at the protein level, Western blot analysis was performed in parental K562 cells and TKI-resistant derivatives after STAT5A targeting. As shown in Fig. 4A, STAT5A-knockout cells exhibited a marked reduction in STAT5A protein expression compared to non-targeting control groups in all cell models. Densitometric analysis (Fig. 4B) further confirmed a consistent decrease in STAT5A protein levels, demonstrating the functional efficiency of the CRISPR/Cas9 editing strategy. β-actin was used as a loading control to ensure equal protein loading across samples. These results indicate that the observed phenotypic and transcriptional effects in subsequent experiments are associated with effective STAT5A depletion.
Fig. 4 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Validation of CRISPR/Cas9-mediated STAT5A knockout at the protein level in K562 cells. A Representative Western blot analysis showing STAT5A protein expression (~ 90 kDa) in untreated (UT) K562 cells, imatinib-resistant (1.2 µM Ima/Res UT), ponatinib-resistant (3 nM Pon/Res UT), STAT5A-knockout (STAT5A KO), and STAT5A-knockout resistant cell lines. β-actin (~ 41 kDa) was used as a loading control. B Densitometric quantification of STAT5A protein levels normalized to β-actin and expressed as fold change relative to untreated control cells. Data represent mean ± SD from three independent experiments. Statistical significance was determined using one-way ANOVA followed by Tukey’s post hoc test (*p < 0.05)
CRISPR/Cas9-mediated STAT5A knockout induces apoptosis in K562 and TKI-resistant cellsSTAT5A was knocked out in both K562 control cells and resistant K562 cell lines using CRISPR/Cas9, and apoptotic effects were analyzed 48 h after transfection. In K562 control cells, the apoptosis rate was 9.9% early apoptosis, 3.1% late apoptosis, and 0.8% necrosis, while knocking out STAT5A in K562 cells resulted in 17.7% early apoptosis, 7.5% late apoptosis, and 1.6% necrosis, representing a 1.93-fold increase in apoptosis compared to the control group. In the 1.2 µM imatinib-resistant K562 cell line, STAT5A knockout induced 16.3% early apoptosis, 6.4% late apoptosis, and 0.9% necrosis, corresponding to an 11.35-fold increase in apoptosis compared to the control group. In the 3 nM ponatinib-resistant K562 cells, the control group exhibited 8.4% early apoptosis, 0.6% late apoptosis, and 0.2% necrosis. Knockout of STAT5A increased these rates to 22.3% early apoptosis, 14.4% late apoptosis, and 5% necrosis, a 4.07-fold increase in apoptosis compared to the control group. These results indicate that knockout of STAT5A significantly increases apoptotic processes in both the control group and the resistant K562 cell lines (Fig. 5). This apoptotic enhancement was statistically significant across all biological replicates, supporting the robust pro-apoptotic effect of STAT5A depletion.
Fig. 5 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.A Apoptosis was analyzed 48 h after transfection in K562 control cells, K562 TKI-resistant control cells, and in groups treated with CRISPR/Cas9 for STAT5A knockout (B) Fold change in apoptosis levels measured 48 h after transfection in K562 control and STAT5A knockout (KO) cells (mean ± SD 1.93 ± 0.075, P = 0.0005). C Fold change in apoptosis levels measured 48 h after transfection in K562/1.2 μm Imatinib resistant cells and K562/1.2uMimatinib resistant STAT5A knockout (KO) cells (mean ± SD 11.53 ± 0.66, P = 0.011) (D) Fold change in apoptosis levels measured 48 h after transfection in K562/3nM ponatinib resistant cells and K562/3nM ponatinib resistant STAT5A knockout (KO) cells (mean ± SD 4.07 ± 0.26, P = 0.0014). Statistical significance was determined using an unpaired two-tailed Student’s t-test
STAT5A knockout modulates cell cycle progression in K562 and TKI-resistant K562 cell linesIn the K562 control cells and the resistant K562 cell lines, STAT5A was knocked out using CRISPR/Cas9, and cell cycle analysis was performed 48 h after transfection. In K562 control cells, the distribution of cell cycle phases was 46.3% G0/G1, 25.2% S, and 19.2% G2/M. After knockout of STAT5A, K562 cells exhibited 43.5% G0/G1, 27.5% S, and 19.9% G2/M, corresponding to a 0.93-fold decrease in G0/G1 and a 1.07-fold increase in S phase compared to control (Fig. 5A-B). In 1.2 µM imatinib-resistant K562 cells, the control distribution was 41.8% G0/G1, 36.9% S, and 21.7% G2/M, while STAT5A knockout resulted in 43.4% G0/G1, 34.3% S, and 21.3% G2/M, corresponding to a 1.04-fold increase in G0/G1 and a 0.95-fold decrease in S phase compared to control (Fig. 6A-C). In 3 nM ponatinib-resistant K562 cells, control cells exhibited 46.0% G0/G1, 24.3% S, and 24.7% G2/M. Knockout of STAT5A resulted in 50.4% G0/G1, 23.5% S, and 19.4% G2/M, corresponding to a 1.09-fold increase in G0/G1 and a 0.78-fold decrease in G2/M compared to control (Fig. 6A-D).These results indicate that knockout of STAT5A affects cell cycle progression in both the K562 control and the imatinib- and ponatinib-resistant cell lines, primarily through an increase in G0/G1 phase and changes in S and G2/M phases (Fig. 6).
Fig. 6 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.A Cell cycle analysis performed 48 h after transfection in K562 control, imatinib-resistant, and ponatinib-resistant cells, as well as in K562 STAT5A knockout and resistant STAT5A knockout cell lines. B Fold change graph of cell cycle analysis in K562 control and K562 STAT5A knockout cells (G0/G1 mean ± SD = 0.93 ± 0.095, P = 0.0034, S: mean ± SD = 1.07 ± 0.060, P = 0.0010, G2/M mean ± SD = 1.03 ± 0.02, P = 0.0001) (C) Fold change graph of cell cycle analysis in K562/1.2 μm imatinib resistant cells and K562/1.2 μm imatinib resistant STAT5A knockout cells (G0/G1 mean ± SD = 1.03 ± 0.085, P = 0.0022, S: mean ± SD= 0.94 ± 0.15, P = 0.0082, G2/M mean ± SD = 0.98 ± 0.05, P = 0.0009) (D) Fold change graph of cell cycle analysis in K562/3nM ponatinib resistant cells and K562/3nM ponatinib resistant STAT5A knockout cells (G0/G1 mean ± SD = 1.09 ± 0.04, P = 0.0004, S: mean ± SD= 0.96 ± 0.13, P = 0.0060, G2/M mean ± SD = 0.78 ± 0.19, P = 0.0189) Statistical significance was determined using an unpaired two-tailed Student’s t-test
STAT5A knockout modulates the expression of apoptosis and survival genes in K562 and TKI-resistant cellsFollowing STAT5A knockout, RT-qPCR profiling revealed distinct yet interconnected transcriptional alterations across parental and TKI-resistant K562 models (Fig. 7A–C), underscoring the context-dependent regulatory functions of STAT5A in chronic myeloid leukemia. In parental K562 cells, deletion of STAT5A led to a pronounced repression of STAT3 (−10.61)—a central mediator of survival and proliferation—accompanied by downregulation of BAX (−0.93) and RUNX3 (−2.48), suggesting suppression of both intrinsic apoptotic triggers and tumor-suppressive transcriptional programs. In parallel, ATM was markedly upregulated (2.92), reflecting activation of DNA damage response signaling as a compensatory mechanism, while BCR (0.75) modestly increased, possibly indicative of feedback activation within BCR::ABL1–dependent circuits (Fig. 7B).
In imatinib-resistant K562 cells (1.2 µM), STAT5A loss reshaped the transcriptome toward an apoptotic and stress-responsive state, evidenced by the upregulation of TP53 (1.50), BCL-X (0.97), CASP3 (1.15), CISH (1.16), and mTOR (1.40) (Fig. 7B). These changes highlight a dual effect: (i) reactivation of p53-driven apoptotic cascades, and (ii) partial engagement of adaptive mTOR and CISH signaling, likely functioning as compensatory feedback loops within the JAK/STAT axis. Meanwhile, STAT5B (1.13) upregulation denotes a possible isoform-level compensation for STAT5A loss, although CASP8 (−1.64) and ATM (−1.95) downregulation suggests reduced extrinsic apoptotic priming and dampened DNA repair signaling under imatinib stress. In ponatinib-resistant cells (3 nM), STAT5A knockout elicited a distinctive pro-apoptotic signature characterized by significant activation of CASP8 (2.24), BCL-X (1.53), BCR (2.06), and TP53 (1.04), coupled with notable repression of STAT5B (−1.57) and JAK2 (−0.83) (Fig. 7C). These alterations collectively imply that STAT5A ablation in ponatinib-resistant cells enhances extrinsic apoptosis via CASP8 activation, while concurrently attenuating JAK2/STAT5B–mediated survival signaling, effectively disrupting compensatory resistance pathways.
Collectively, these transcriptional landscapes reveal that STAT5A acts as a master node orchestrating survival–apoptosis balance. In addition, STRING-based protein–protein interaction analysis revealed dense connectivity among STAT5A, TP53, CASP3, and STAT3, reinforcing their cooperative role in modulating apoptotic and DNA damage responses. Its disruption reprograms resistant CML cells toward apoptotic sensitivity and reduced proliferative capacity. Importantly, the magnitude and directionality of these changes are context-specific: STAT3 repression and ATM activation dominate in parental cells, TP53 and CASP3/BCL-X reactivation define the imatinib-resistant profile, and CASP8-driven extrinsic apoptosis underlies the ponatinib-resistant response. Thus, STAT5A knockout reinstates apoptotic competency through both intrinsic and extrinsic pathways, dismantling key resistance circuits and positioning STAT5A as a therapeutically actionable vulnerability in TKI-refractory CML.
STAT5A knockout reveals key transcriptional changes in control and TKI-resistant K562 cellsIn untreated K562 control cells (K562 UT), absolute expression values were generally low across most genes, except for BAX (2^-ΔCt = 0.23) and RUNX3, which showed relatively higher basal expression (Fig. 7a). Following STAT5A knockout, expression of pro-apoptotic genes BAX (0.23 → 0.12) and CASP3 (0.08 → 0.04) decreased, while CASP8 remained unchanged, suggesting a mild suppression of intrinsic apoptotic signaling. Anti-apoptotic markers BCL-X and BCL2 showed minimal variation. Meanwhile, STAT1 and STAT3 levels slightly declined, aligning with attenuation of survival signaling. DNA repair and proliferation-associated genes ATM and ABL1 were notably downregulated, whereas TP53, mTOR, and RUNX3 showed modest decreases (Figs. 6A and 7B). These data suggest that STAT5A ablation in parental K562 cells induces a restrained apoptotic response accompanied by impaired proliferative signaling.
In imatinib-resistant K562 cells (1.2 µM Ima/Res UT), baseline expression levels of STAT5A, BCL-X, and CASP3 were slightly elevated compared to control, consistent with a survival-oriented transcriptional state (Fig. 7A). Upon STAT5A knockout, expression of TP53, BCL-X, CASP3, and RUNX3 increased, reflecting partial reactivation of apoptotic and tumor-suppressive pathways (Fig. 7B and C). Concomitantly, upregulation of STAT1, mTOR, and CISH suggested activation of compensatory feedback circuits within the JAK/STAT axis, while CASP8 and ATM suppression implied reduced extrinsic apoptosis and DNA repair signaling. Collectively, these shifts indicate a dual remodeling effect: (i) reactivation of p53-driven apoptotic cascades and (ii) adaptive signaling realignment to counter imatinib-induced stress.
Fig. 7 The alternative text for this image may have been generated using AI.
The alternative text for this image may have been generated using AI.Transcriptional reprogramming following STAT5A knockout in K562 and TKI-resistant models. A Comparative log₂ fold change of selected genes across baseline, imatinib-resistant (1.2 µM), and ponatinib (3 nM) conditions in STAT5A-knockout K562 cells (dashed line = 0). B Differential expression (log₂ fold change) of apoptosis- and signaling-related genes in STAT5A-knockout K562 cells under baseline and imatinib-resistant (1.2 µM) conditions (dashed line = 0). C Differential expression (log₂ fold change) of selected genes in STAT5A-knockout K562 cells under baseline and ponatinib (3 nM) conditions (dashed line = 0)
In ponatinib-resistant K562 cells (3 nM Pon/Res UT), STAT5A and STAT3 expression markedly increased (2^-ΔCt = 1.31 and + 0.83), consistent with strong survival signaling (Fig. 8A). After STAT5A deletion, a distinct transcriptional rewiring occurred—CASP8, BCL-X, and TP53 were upregulated, while STAT5B and JAK2 were repressed (Figs. 6C and 7D). These alterations highlight a context-specific apoptotic re-sensitization, in which extrinsic apoptosis via CASP8 activation is restored, and JAK2/STAT5B-mediated survival is attenuated. The concomitant downregulation of STAT3 and ATM further suggests disruption of DNA repair and proliferative control mechanisms. When integrated across all models (Figs. 7A and C and 8A and D), STAT5A knockout consistently reprogrammed transcriptional networks toward apoptotic activation and stress adaptation in a resistance-dependent manner. In parental cells, repression of STAT3 and induction of TP53/RUNX3 emphasize disruption of proliferative control and restoration of tumor-suppressive signaling. In imatinib-resistant cells, activation of CASP3, STAT1, and mTOR denotes an apoptotic shift coupled with metabolic rewiring. Finally in ponatinib-resistant cells, CASP8- and TP53-driven apoptosis was reestablished, undermining key resistance circuits. Collectively, these findings establish STAT5A as a transcriptional hub balancing survival and apoptosis. Its loss dismantles STAT3/STAT5B-dependent survival pathways, reinstates intrinsic (BAX, CASP3, TP53) and extrinsic (CASP8, BCL-X) apoptotic circuits, and restores apoptotic competency across resistant states, underscoring its therapeutic vulnerability in TKI-refractory CML.
Together, these results demonstrate that STAT5A depletion reprograms transcriptional and phenotypic outcomes across both parental and TKI-resistant CML models. By reinstating apoptotic competence and disrupting compensatory STAT3/STAT5B signaling, STAT5A knockout effectively restores TKI sensitivity. The following Discussion contextualizes these findings within existing literature and explores their translational implications for CML therapy.
Comments (0)