
Remember me
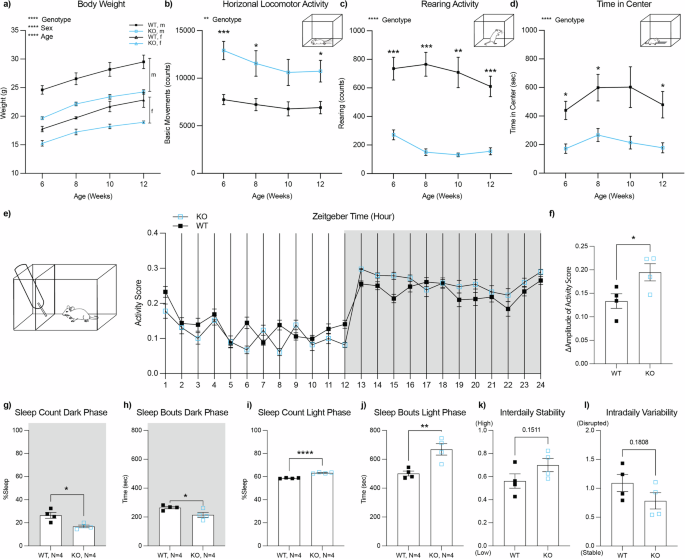

Compared to WT controls, Adgrl3 KO mice weighed less (Figs. 1a and S1A, E) and travelled greater horizontal distances along both X and Y axes in the open field test (Figs. 1b and S1B, F; S2A, B), indicating increased baseline locomotor activity, consistent with previous reports in this model [14, 16]. They also showed reduced rearing behavior (Figs. 1c and S1C, G) and spent less time in the center of the enclosure (Figs. 1d and S1D, H), a pattern confirmed by spatial distribution analysis (Fig. S2C, D). While reduced center time and rearing are often interpreted as anxiety-like behavior [25], these measures may be confounded by increased activity in Adgrl3 KO mice.
Fig. 1: Adgrl3 knockout mice show increased activity and altered sleep patterns.
a Body weight of wild-type (WT) and Adgrl3 knockout (KO) mice of both sexes (WT: n = 9; 5 males, 4 females; KO: n = 17, 13 males, 4 females). Three-way ANOVA was performed to analyze the effect of age, sex, and genotype on body weight (****, p < 0.0001). b–d Horizontal locomotor activity, rearing, and center time across development. Two-way repeated measures ANOVA was performed to analyze the effect of age and genotype on each dependent variable (basic movements, rearing, or time in center). This was followed by Ṥídák’s multiple comparisons test (*, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001). e, f PiezoSleep system activity scores and amplitude across Zeitgeber time (WT: n = 4; 1 male, 3 females; KO: n = 4, 1 male, 3 females). g–j Sleep metrics during dark and light cycles: percent time asleep and average bout duration. k, l Interdaily stability and intradaily variability quantify rhythm regularity and fragmentation [49]. Unpaired t-tests used for all PiezoSleep comparisons (****, p < 0.0001; ***, p < 0.001; **, p < 0.01; and *, p < 0.05). Illustrations used in this image were adapted from SciDraw.
Adgrl3 knockout mice exhibit increased activity during the dark cycle compared to controlsTo assess whether elevated activity in Adgrl3 KO mice varies by time of day, we monitored sleep/wake behavior using the PiezoSleep system over five days under a 12-hour light/dark cycle. Adgrl3 KO mice showed elevated activity across the light-dark transition (Fig. 1e, f), likely driven by reduced sleep and shorter sleep bouts during the dark (active) phase (Fig. 1g, h). This is consistent with prior findings in Adgrl3 KO rats [17]. During the light cycle, Adgrl3 KO mice slept more and had longer sleep bouts (Fig. 1i, j), suggesting an altered sleep-wake regulation. We also analyzed interdaily stability, a measure of daily consistency, and intradaily variability, which reflects fragmentation of activity within a 24-hour period (Fig. 1k, l). Neither measure differed significantly between genotypes.
Adgrl3 KO mice show increased licking behavior for evaporated milkTo assess hedonic drive, WT and Adgrl3 KO mice were tested in a six-bottle preference task using a gustometer (Fig. S3) [26]. While previous studies in Adgrl3 KO mice have examined motivational responses to food rewards, the hedonic (“liking”) component has not been directly assessed. Mice sampled water and five concentrations of evaporated milk (6.25–100%) over 30 min. Adgrl3 KO mice completed more trials, initiated licking more frequently, and showed shorter latencies to first lick compared to WT controls (Fig. S3B–E), suggesting enhanced reward engagement. However, when analysis was restricted to completed trials, the average lick count per solution did not differ between genotypes (Fig. S3F–H), suggesting that the elevated responding in Adgrl3 KO mice reflects increased instrumental engagement rather than enhanced hedonic “liking” per se. Fully dissociating these components will require additional paradigms, such as consumption measures under conditions that minimize instrumental demand, and remains a question for future work.
Adgrl3 KO mice have greater levels of evoked dopamine release in the striatumWe used Fast-Scan Cyclic Voltammetry (FSCV) to detect electrically evoked dopamine with sub-second precision in brain slices of the dorsal and ventral striatum of adult mice [27] (Fig. 2). Compared to WT controls, Adgrl3 KO mice showed significantly higher levels of evoked dopamine release in both the dorsal (Fig. 2b, d) and ventral striatum (Fig. 2f, h). Despite the elevated response, there were no significant genotype differences in dopamine reuptake in either region, as indicated by the rate of dopamine signal decay (Tau1) following stimulation (Fig. 2c, g).
Fig. 2: Adgrl3 KO mice have greater levels of evoked dopamine (DA) in the striatum.
Coronal sections that included striatum were obtained from wild-type (WT) and Adgrl3 knockout (KO) mice aged P102-P183. a Schematic of location for fast-scan cyclic voltammetry (FSCV) recordings from the dorsal striatum. Slices were stimulated with a bipolar concentric electrode and DA was detected using a carbon fiber electrode. b FSCV recordings of evoked DA release from the dorsal striatum (N = 6 WT, N = 8 KO; 31 slices WT, 44 slices KO). c DA reuptake from the dorsal striatum (N = 5 WT, N = 7 KO; 25 slices WT, 47 slices KO). d Averaged dopamine transients in response to a single electrical pulse in the dorsal striatum from all tested mice, illustrating signal kinetics. e Schematic of location for FSCV recordings from the ventral striatum. f FSCV recordings of evoked DA release from the ventral striatum (N = 5 WT, N = 7 KO; 24 slices WT, 36 slices KO). g DA reuptake from the ventral striatum (N = 4 WT, N = 7 KO; 19 slices WT, 42 slices KO). h Averaged dopamine transients in response to a single electrical pulse in the ventral striatum from all tested mice, illustrating signal kinetics. All quantal release events were analyzed in IgorPro using a publicly available GitHub-based analysis program [50]. Peak parameters included amplitude (Imax, pA) and event duration (Tau1, ms). All statistical analyses were performed using an unpaired t-test (*, p < 0.05). Illustrations used in this image were adapted from SciDraw.
Dopamine release during a continuous reinforcement task is not significantly altered in Adgrl3 KO miceIn the nucleus accumbens, dopamine is released in response to unexpected rewards [20, 28]. With training, this signal shifts to predictive cues. We hypothesized that Adgrl3 KO mice would exhibit enhanced cue-induced dopamine release based on prior studies in brain slices [18] and our own ex vivo FSCV data. To test this, we used in vivo fiber photometry with dLight1.2 [29] during a continuous reinforcement (CRF) task (Fig. 3).
Fig. 3: Performance of WT and Adgrl3 KO mice in a continuous reinforcement task (CRF).
a (Left) Experimental timeline illustrating progression from viral injection and optic fiber implantation to fiber photometry. (Middle) Optic fibers were unilaterally implanted into the nucleus accumbens. A representative coronal section shows dLight1.2 expression in the striatum and the location of the optic fiber, with dashed lines indicating the fiber shaft. (Right) WT and Adgrl3 knockout (KO) mice were trained on a fixed-ratio 1 schedule of reinforcement, in which each lever press was rewarded with evaporated milk from a dipper. Training occurred over seven sessions on separate days. b Percentage of lever presses relative to total possible lever presentations. c Percentage of trials in which mice lever pressed and consumed the evaporated milk reward. Statistical analysis in (b) and c consisted of a two-way ANOVA followed by Ṥídák’s multiple comparison test (*, p < 0.05; **, p < 0.01; ***, p < 0.001; ****, p < 0.0001). (N = 14 WT, 13 KO). d Following training, WT and Adgrl3 KO mice completed a continuous reinforcement task where they could earn up to 60 rewards per session. Dopamine levels were monitored via fiber photometry during this operant task. e Average dLight1.2 traces aligned to lever extension for WT and Adgrl3 KO mice. Shaded area (yellow) indicates the timeframe used for AUC calculation. f Area under the curve (AUC) and g peak height for the dLight1.2 traces. Statistical analysis consisted of an unpaired t test. (N = 9 WT, 10 KO). Note: Some animals were excluded from photometry due to inadequate fiber placement or insufficient signal. Illustrations used in this image were adapted from SciDraw.
After viral injection and fiber implantation (Fig. S4), mice were trained to retrieve evaporated milk rewards from a dipper port (30 “free” presentations per session). We found no significant difference in the percentage of rewards retrieved or the total number of rewarded and non-reward head entries between WT and Adgrl3 KO mice (Fig. S5).
Mice were then trained on a lever-pressing task with a fixed-ratio 1 schedule, where each lever press triggered a reward. Adgrl3 KO mice acquired lever pressing behavior more quickly than WT mice, completing ~90% of trials in the first session compared to ~22% in controls (Fig. 3b). To determine whether KO mice associated lever pressing with receiving the evaporated milk reward, we measured the percentage of trials in which mice both pressed the lever and consumed the reward (Fig. 3c). In the first session, WT and KO mice had the same percentage of rewarded trials, indicating that despite engaging in more lever pressing, Adgrl3 KO mice did not exhibit a stronger association between lever pressing and reward. By session four, Adgrl3 KO mice reached near-perfect performance, indicating faster acquisition of the cue-reward association compared to WT mice, which required seven sessions to reach similar levels.
To further assess learning dynamics, we analyzed lever press and reward latencies across training sessions. Adgrl3 KO mice pressed the lever significantly faster in early sessions, but when matched by performance level ( ≥ 50% trial completion), latencies were comparable across genotypes. Both groups improved over time, reaching ~5 s for lever press latency and ~1 second for reward retrieval latency (Fig. S6A, B).
Following training, mice completed three CRF sessions where they could earn up to 60 rewards per session. Both genotypes completed all trials and reduced session duration over time (Fig. 3d). During these sessions, dopamine release aligned to lever extension was measured using dLight1.2. No statistically significant differences were observed in either the area under the curve (AUC) or peak amplitude between genotypes (Fig. 3e–g).
Finally, we examined the relationship between dopamine release and lever press latency. Across genotypes, greater dopamine release was associated with faster lever pressing, and correlation strength did not differ between groups (Fig. S7).
Fixed interval task reveals longer latencies and reduced dopamine release in Adgrl3 knockout miceTo further investigate dopamine release during behavior, we tested mice in a fixed interval (FI) operant task, which requires waiting a set duration before a lever press will trigger reward delivery (Fig. 4a). Unlike CRF, where each lever press immediately and predictably delivers reward, the FI task dissociates the cue from reward delivery by varying the interval between them, making it a more sensitive assay for detecting how dopamine signals scale with changing cue-reward contingencies. Mice were tested across five fixed intervals (2, 4, 8, 12, and 24 s), with three sessions per interval. During each trial, the lever was extended, but only the first press after the designated interval resulted in a reward.
Fig. 4: Performance of WT and Adgrl3 KO mice in a fixed interval task.
a Mice were tested across five fixed intervals (2-24 s), with three sessions per interval. During each trial, the lever was extended for the designated interval, after which it remained extended until pressed. A successful press delivered a 10-second evaporated milk reward. Each session included up to 60 rewarded trials, with a variable intertrial interval. b Total lever presses per interval. No significant genotype differences. (N = 10 WT, 10 KO). c Latency to first lever press, d average latency to lever press, and e latency to reward across sessions. All statistical analyses were performed using a two-way ANOVA with Ṥídák’s multiple comparison test (**, p < 0.01; ***, p < 0.001; ****, p < 0.0001). (N = 8 WT, 10 KO). f, g Average dLight1.2 traces aligned to lever extension for WT and Adgrl3 KO mice. h Area under the curve (AUC) and peak height from 0-1.5 s. Statistical analysis was performed using a two-way ANOVA with Ṥídák’s multiple comparison test (*, p < 0.05; **, p < 0.01; ***, p < 0.001). (N = 8 WT, 10 KO). i, j Average dLight1.2 traces aligned to dipper extension for WT and Adgrl3 KO mice. k AUC and peak height from 0-1.0 s. Statistical analysis was performed using a two-way ANOVA followed by Ṥídák’s multiple comparison test (**, p < 0.01; ****, p < 0.0001). (N = 8 WT, 10 KO). Note: Some animals were excluded from photometry due to inadequate fiber placement or insufficient signal. Illustrations used in this image were adapted from SciDraw.
Both genotypes increased lever pressing as the interval lengthened and exhibited the expected “scalloping” pattern within sessions, where response rates gradually increased as the time for the next reward approached (Fig. S8). This pattern, along with the similar total lever press counts between groups (Fig. 4b), suggests that temporal processing of the interval structure was largely intact in Adgrl3 KO mice.
Significant genotype differences emerged in latency measures. Adgrl3 KO mice took longer to initiate lever pressing across intervals (Fig. 4c) and showed increased average press latency across sessions (Fig. 4d). Both groups, however, displayed similar patterns across intervals, indicating that these differences are not due to impaired adaptation to the changing contingencies. KO mice also exhibited longer reward-retrieval latencies (Fig. 4e). These results suggest reduced task engagement or delayed response execution.
We next examined dopamine release during the FI task using dLight1.2 fiber photometry. When aligned to lever extension, dopamine signals decreased with increasing interval length, consistent with reduced reward certainty. WT mice showed higher dopamine levels at shorter intervals and a larger drop across intervals (ΔAUC = 68 units), whereas KO mice started lower and showed a smaller decrease (ΔAUC = 39 units) (Fig. 4f, g). We next aligned the signal traces to dipper extension, defined as the moment of reward delivery. Both genotypes showed increasing dopamine responses as the fixed interval and the delays between cue and reward lengthened. However, WT mice showed a larger increase (ΔAUC = 82 units) compared to Adgrl3 KO mice (ΔAUC = 14.5 units) (Fig. 4i–k). Together, these findings show that Adgrl3 KO mice fail to scale cue-evoked dopamine signals to task contingencies, despite intact interval-dependent changes in press latencies (Fig. 4c, d).
Amphetamine challenge suggests comparable dopamine release capacity in WT and Adgrl3 KO miceOur ex vivo FSCV findings indicated increased dopamine release capacity in KO mice (Fig. 2), yet in vivo photometry revealed reduced cue-evoked dopamine. To determine whether release capacity is altered in vivo, we administered increasing doses of amphetamine (1.5, 5, 10, and 20 mg/kg, s.c.) and monitored dopamine dynamics using fiber photometry (Fig. 5). Locomotor activity was simultaneously tracked in an open field using AnyMaze.
Fig. 5: Open field test with amphetamine challenge suggests phasic dopamine release capacity is similar between WT and Adgrl3 KO mice.
a Mice injected with AAV5-hSyn-dLight1.2 and implanted with an optic fiber in the nucleus accumbens were used to measure dopamine responses to amphetamine in an open field test. Following a 30-min habituation period, mice received a saline injection to assess the effects of injection stress on dopamine signaling, followed by amphetamine administration (1.5, 5, 10, or 20 mg/kg subcutaneous) on separate days. Dopamine activity was recorded using fiber photometry, and locomotor behavior was tracked with AnyMaze software (Stoelting Co., Wood Dale, IL). b Difference in average distance traveled by WT and Adgrl3 KO mice injected with saline versus varying doses of amphetamine. Statistical analysis was performed using a two-way ANOVA followed by Ṥídák’s multiple comparison test (**, p < 0.01). ****, p < 0.0001 for genotype and ***, p < 0.001 for dose. (N = 5 WT, 5 KO). Some data points were excluded due to inadequate video analysis. Average dLight1.2 traces aligned to amphetamine injection for c WT and d Adgrl3 KO mice. e Mean dF/F, f slope, and g latency to reach plateau for WT and Adgrl3 KO mice following amphetamine injection at the four doses. Statistical analysis was performed using a two-way ANOVA (*, p < 0.05; **, p < 0.01; ****, p < 0.0001). (N = 5 WT, 5 KO). Illustrations used in this image were adapted from SciDraw.
In WT mice, 5 mg/kg and 10 mg/kg doses increased locomotion, while 20 mg/kg reduced activity, consistent with well-established stereotypy observed at high doses [30]. In contrast, Adgrl3 KO mice showed reduced locomotor activity across all doses (Fig. 5b), suggesting altered behavioral sensitivity to amphetamine.
Despite these behavioral differences, dopamine signal traces aligned to amphetamine injection revealed no significant genotype differences in mean dFF, slope, or latency to plateau (Fig. 5c, d). These data indicate that dopamine release capacity is comparable between genotypes and rule out sensor saturation as an explanation for the reduced dopamine signals observed during the FI task.
We also analyzed spontaneous dopamine release (Fig. S9). The total number of spontaneous events did not differ significantly between WT and Adgrl3 KO mice. Event frequency decreased with amphetamine in both genotypes, especially at higher doses. Event duration increased dose-dependently, but peak amplitude remained unchanged.
Comments (0)