
Remember me
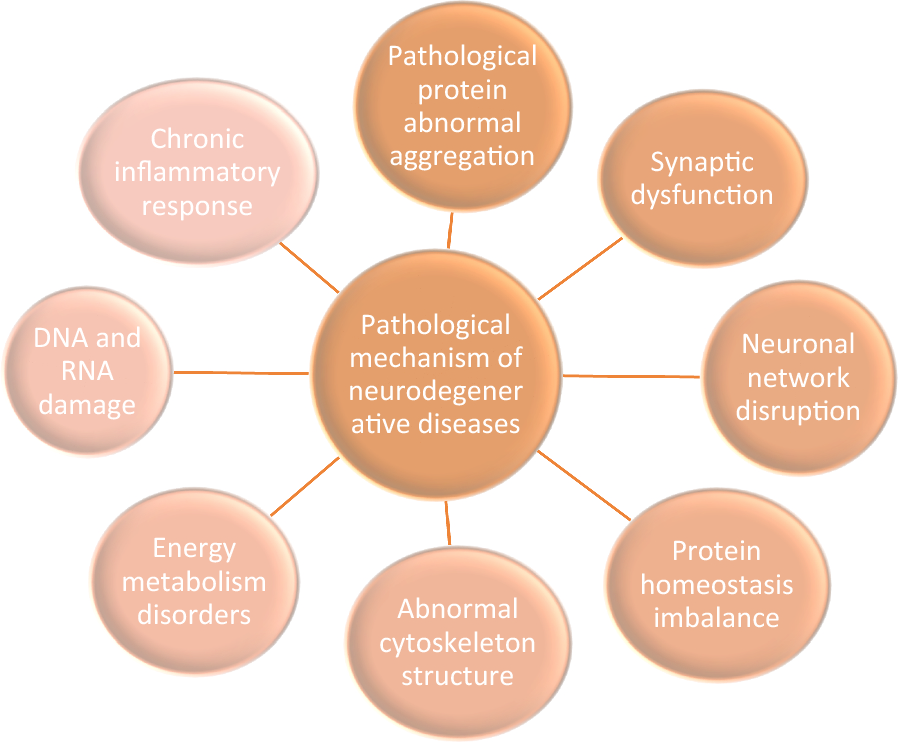

Large animal models, such as primates, provide unique insights into the pathogenesis of NDs, which are not always evident in rodent models. Compared with rodents, primates have differences in the expression and function of proteins such as PINK1 in their brains. Mutations in the human PINK1 and PRKN genes can easily lead to neurodegeneration and mitochondrial dysfunction, resulting in Parkinson’s disease. Genetic based amyloid pathology models in rodents may not fully replicate the complexity of human NDs. Larger animal models can be closer to human responses in order to understand the impact of EE [110]. A study using CRISPR/Cas9 to target large animals such as pigs and monkeys has found pathological events similar to neurodegeneration in patients’ brains, which cannot be replicated in small animal models. The genome editing of non-human primates is performed by introducing the CRISPR/Cas9 system into fertilized eggs. Although large animals have unique properties for modeling NDs, there are also limitations in testing treatment efficacy [111]. Moreover, EE research has major limitations such as inter laboratory variability, difficulty in separating different components of EE, and differences in individual responses to EE [112]. EE may trigger neuroprotective effects as well as neurogenic transcription and translation events [27]. In various neurodegenerative disease mouse models, specific changes targeting the underlying pathological mechanisms of the diseases can be observed.
Enriched environment therapy in neurodegenerationChallenges in translating animal models to humansOne of the main challenges in translating research results from animal models into humans is the variability of responses due to species-specific biological differences [113]. Although EE has shown promising results in enhancing neurogenesis and cognitive function in animal studies, the same effect may not be as apparent in humans due to differences in brain structure and function. In addition, ethical considerations and the complexity of human diseases pose significant obstacles to effective translation. In the future, it is necessary to develop large-scale animal models that are closer to human anatomy and physiology, as well as integrate advanced technologies such as imaging and biomarker analysis, to improve the predictive effectiveness of animal research [114].
In AD research, the 5xFAD mouse model emulated the pathological hallmarks of the disease, including β-amyloid plaques and tau protein neurofibrillary tangles, by expressing human genes harboring mutations associated with familial Alzheimer’s disease [27, 115]. Cognitive function in these mice was evaluated using behavioral assays such as the Morris water maze, which measured spatial memory and learning capabilities. Pathological alterations were quantified through immunohistochemical and Western blot analyses [27]. For PD, the MPTP-induced mouse model simulated the disease’s pathological characteristics by selectively targeting dopaminergic neurons. Motor function was assessed using the rotarod test and the pole test, which evaluated balance, strength, coordination, and bradykinesia, respectively. And the number of dopaminergic neurons in the substantia nigra was determined using immunohistochemical quantification [116]. In amyotrophic lateral sclerosis study, the SOD1 mutant mouse model replicated the disease’s pathological features, such as the degeneration of motor neurons, by expressing mutated human SOD1 genes. Muscle strength was assessed through grip strength tests, while the health of motor neurons was indicated by nerve conduction velocity, as evaluated by neurophysiological testing [117]. In recent decades, significant progress has been made in PD modeling, but the elderly population still lacks effective treatment for PD. The PD model, from cellular to toxicity and genetic animal models, replicated almost all the features of the disease and was capable of identifying specific events and testing new neuroprotective strategies. Classic and new models effectively supplemented PD feature modeling and enhanced current understanding of disease molecular mechanisms [118].
In the R6/1 HD mouse model, EE may promote the recovery of brain-derived neurotrophic factor transport in the striatum. In Alzheimer’s disease models, EE may enhance the levels of the amyloid-degrading enzyme Carboxypeptidase E (CPE) [119]. These findings reveal the potential role of EE in modulating microglial responses and inflammation, potentially providing new therapeutic targets for the intervention of NDs in the future [50, 120].
Traditionally, rodent models have been the cornerstone of neurodegenerative disease research, but they still have certain limitations in summarizing human brain structure and disease pathology. Therefore, some people also use large animal models for study [111]. The CRISPR/Cas9 gene editing technology provides a foundation for the creation of large animal models, such as pigs and monkeys, which are more similar to human neuroanatomical and physiological features. A Huntington’s disease pig model has been developed that exhibits selective neurodegeneration and motor dysfunction similar to human HD patients [111]. The latest progress also emphasizes the role of fruit fly models in drug discovery for NDs. Drosophila provides a genetically tractable system with clear neuroanatomical structures that can quickly identify new therapeutic targets and evaluate drug efficacy. The key molecular pathways between fruit flies and humans are highly conserved, making the simulation of human NDs more reliable [121].
Influences of environment and individuals on enriched environmentIn the study of EE, potential confounding factors such as variations in environmental conditions and individual differences in animals can lead to different effects of EE therapy. The same differences also exist in human. A study found a causal relationship between exposure to common environmental factors and major NDs, and multiple environmental factors have overlapping effects on NDs [122]. Variations of environmental conditions such as environmental toxins can also increase the risk of developing major NDs. Neurotoxic metals can produce senile plaques or amyloid plaques and neurofibrillary tangles, leading to neurological dysfunction [123].
The effect of EE on NDs may vary depending on the gender, genetic background, or environmental factors of the subjects [124]. Studies have shown that most of the clinical differences reported in NDs appeared to be closely related to gender, which might be evidenced by significant changes in female/male autoimmune mechanisms, reflecting the impact of gender differences on EE efficacy [125]. The impact of social and physical EE on brain plasticity and cognition varied, resulting in different outcomes due to the types of environmental enrichment that affected disease efficacy [125].
Adult neurogenesis is a potential target for extending cognitive health lifespan, but it decreases with age. Based on this, EE may be more effective in the early stages of NDs before significant cognitive decline [126]. There has also been study discussing that the complexity of electroencephalogram (EEG) signals might be influenced by EE, showing a U-shaped pattern with age, increasing from late adolescence to adulthood and decreasing in old age. This pattern suggests that EE may have different impacts throughout the entire lifecycle, and middle-aged individuals may be more effective compared to older age [127]. Considering the progression stage of the disease, EE is generally more effective in the preclinical stage of NDs [128].
Epigenetic modifications enriched by environmental influences can promote long-term neuroprotection and disease progression in NDs. There are differences in epigenetic modifications between individuals, which may affect the impact of NDs on individuals and lead to differences in the response of different individuals to EE [129]. A study found that the intrinsic activity intensity of brain functional networks had a high heritability, while the functional connections between brain networks were influenced by environmental factors. This indicated that in EE research, an individual’s genetic background might affect their response to environmental stimuli, resulting in individual differences [16].
The epigenetic modification mechanisms influenced by EE also vary among different types of diseases. In Alzheimer’s disease, the levels of H3K27ac and H3K9ac histone modifications in the patient’s brain were elevated. These modifications affected the regulation of disease-related genes. The RNA methylation patterns between animal models of Alzheimer’s disease and Parkinson’s disease were also different, indicating that the epigenetic profiles of each disease are different. In addition, elevated DNA methylation could inhibit the expression of genes involved in DNA repair, which was one of the factors contributing to the development of Huntington’s disease [16]. A systematic review indicated that EE could modulate epigenetic processes in the central nervous system under adverse conditions. The benefits of EE on animal brains and behavior are directly linked to distinct epigenetic mechanisms, which are reflected in cellular growth and neuroplasticity. After exposure to EE, an increase in the expression of miR-221 and miR-483 was observed in the prefrontal cortex, while a decrease in the expression of miR-92a-3p and miR-134 was noted in the hippocampus. In terms of DNA modifications, a reduction of DNMT levels in the hippocampus was reported. EE may serve as a non-pharmacological and easily applicable alternative to prevent symptoms affecting brain tissue diseases [130].
Variability and standardization in enriched environment studiesThere is variability and standardization in EE across different studies, and differences in protocol settings can affect the results and reproducibility of experiments. EE components can be variable. The size of the cage, the number of animals raised, the type of experimental animals, the age of the animals at the beginning of EE, the duration of different feedings, the type of control used, the rat strain used, and the gender of the rats may all serve as differences in the experimental protocol. These differences may lead to different behavioral and neurochemical outcomes [16], as the complexity and social interaction opportunities provided by EE can affect cognitive function and neural plasticity.
Different environmental settings have an impact on experimental results, including enhancing synaptic plasticity in the nervous system, improving learning and memory abilities, and slowing down the progression of NDs [76]. Therefore, when designing and conducting EE studies, careful consideration and standardization of environmental settings must be given, including the use of standardized operating procedures between patients and controls, inclusion of the same pre analysis protocol, to ensure the reliability and reproducibility of research results.
The differences in EE schemes and the effects of EE on individuals of different periods or ages can lead to variations in research results. Some studies showed that early postpartum EE could alter the neurodevelopmental program of stress neuroendocrine responses, as well as the volume and morphology of the hippocampus and prefrontal cortex [131]. In addition, EE induced changes in oxidative metabolism capacity and connectivity in brain regions, as measured by cytochrome c oxidase (CCO) activity. There were also studies indicating that anxiety or depression like behaviors decreased and enhanced learning and memory in certain situations, while others reported an increase in anxiety or no significant impact. These differences might be due to variations and changes in the EE plan and period, which could affect the generalizability of the research findings [131].
Synergism of enriched environment with clinical therapiesEE has become a promising non-pharmacological approach that can synergize with other therapeutic methods such as drugs and cell therapies in promoting human brain repair [27]. For instance, studies have manipulated autophagy to investigate how EE influences this process, which is a significant protective mechanism against NDs [132]. Autophagy is an intracellular degradation process that is crucial for maintaining cellular homeostasis. In NDs, disturbances in autophagy may lead to cellular dysfunction and disease. The accumulation of abnormal protein aggregation, as a common cause of NDs, can be reduced through autophagy degradation [133]. Autophagy facilitates the clearance of impaired and senescent organelles, thus preserving cellular equilibrium. The impact of EE on AD was mainly concentrated in the hippocampus and entorhinal cortex, which were associated with memory formation, spatial navigation, and motor behavior. Researchers have developed animal models of NDs, such as Alzheimer’s disease mouse models, and subjected these models to EE [134]. By quantifying the levels of autophagy-associated proteins in the hippocampus, like LC3-II and p62, the effects of EE on autophagy were evaluated. Study indicates that EE can up-regulate the expression of autophagy-related proteins in the hippocampus of rats, and reduce neurodegeneration induced by oxidative stress [35, 134]. Moreover, EE could promote the recovery of limb motor dysfunction after cerebral ischemia through neuroprotection. This recovery might be achieved by regulating the expression of GAP-43, SYN, and Bcl-2/Bax, and depended on neuroprotection of the ischemic penumbra [135].
NDs exhibit heterogeneity in clinical manifestations, with alpha synucleinopathy and tau proteinopathy being the most common disease categories. Due to the different pathophysiological mechanisms of NDs, the application of EE needs to consider disease specificity. For example, in Alzheimer’s disease, EE may play a role by improving spatial learning and memory consolidation, while in Parkinson’s disease, it may affect the course of the disease more by reducing behaviors related to depression and anxiety [99].
EE has been demonstrated to reduce biomarkers of oxidative stress within the rat brain and bolster the efficacy of antioxidant enzymes, suggesting their potential in mitigating oxidative stress-related neuronal damage [136]. This study assessed the effects of EE on oxidative stress through quantification of malondialdehyde (MDA) and superoxide dismutase (SOD) levels within brain tissue, providing insights into their neuroprotective properties. Oxidative stress is a pivotal factor in neuronal damage and demise, prompting examination of the influence of EE on neuronal health and the progression of NDs. Techniques in molecular biology, including Western blot, are employed to evaluate the activation of pathways implicated in antioxidant stress response, particularly focusing on the Nrf2-ARE pathway. Notably, the Nrf2-ARE signaling pathway, when activated, has been shown to ameliorate neural damage induced by oxidative stress, demonstrating its role in neuroprotection [137], and concurrently initiate the NF-κB inflammatory signaling cascade, dampening inflammatory reactions, which may contribute to neurorestoration. The BCSFB serves as a vital protective mechanism within the central nervous system, safeguarding against the infiltration of harmful xenobiotics by preserving barrier integrity. Researchers utilized electron microscopy to assess the effects of EEs on the ultrastructure of the BCSFB and employed immunohistochemical methods to quantify the expression of tight junction proteins, including Occludin, to elucidate the protective mechanisms of environmental factors on the nervous system to delineate the neuroprotective effects of environmental factors and the therapeutic potential of EEs in neurodegenerative disorders.
Additionally, EEs stimulate the Wnt/β-catenin signaling pathway, reinforcing the integrity of the BCSFB and subsequently enhancing its barrier function, which may offer additional neuroprotection [71, 138]. In PD, downregulation of the WβC signaling pathway led to degeneration of dopaminergic neurons, causing motor and cognitive impairments. Pharmacology and stem cell-based therapies that activated WβC signaling improved alpha synuclein accumulation, Lewy activation mutations, and promoted neurogenesis of dopaminergic neurons. In NDs such as HD, ALS, and MS, the WβC signaling pathway was upregulated, but the reasons for this remained to be studied [69].
In examining the role of environmental elements in neurodegenerative disease therapeutics, with a focus on AD, immunohistochemical assays are routinely applied to evaluate the expression levels of Amyloid Precursor Protein (APP) and Aβ plaque deposition, subsequently gauging the influence of EEs on AD’s pathological hallmarks [139]. By employing this evaluative approach, one might deduce that EE could diminish Aβ aggregation and augment synaptic plasticity, consequently advancing cognitive performance in murine models of AD [28, 140]. Androgen deprivation therapy (ADT) for treating prostate cancer (PCa) is associated with cognitive impairment, which may exacerbate AD related dementia in elderly men. Androgens may protect cognitive function by inhibiting tau protein phosphorylation. Androgen levels can disrupt the balance of genes sensitive to androgen levels, especially in memory and emotional regions such as the hippocampus and amygdala. This hormone manipulation may lead to long-term cognitive problems and AD through processes such as Aβ aggregation and neurofibrillary tangles (NFT) formation. Animal studies have shown that androgen deprivation can reduce synapses in the hippocampus by 40%, increase the deposition of Aβ, alter neural conduction in the frontal cortex, and impair normal neuronal function [141].
Within the domain of neurovascular restoration, the fluorescent gold neurotracing method is harnessed to track pivotal biomarkers influenced by EE, encompassing the expression levels of Nerve Growth Factor (NGF) and Vascular endothelial growth factor (VEGF). Study showed that EEs facilitate the process of neurogenesis [76, 142]. Using a EE for adjuvant therapy can aid in the reconstruction of nerves and blood vessels, promoting repair after brain injury.
Enriched environment in combination with drugsIn clinical applications, the combination of EE and existing therapies can have greater potential for the treatment of current NDs. Research has shown that stem cell extracellular vesicles have the potential to improve symptoms and quality of life in the clinical treatment of diseases such as Alzheimer’s disease, stroke, and Parkinson’s disease. These extracellular vesicles are rich in neurotrophic factors, which can promote the survival and regeneration of neurons, activate intracellular signaling pathways, promote neuronal growth and synapse formation [143]. Research has shown that EE can promote the generation, stabilization, and strengthening of synapses to alter perceptual and behavioral outputs, which at the level of neuronal characteristics determine whether connections are strengthened and maintained, or eliminated. Early stress conditions, such as maternal care, can lead to long-term emotional and behavioral changes in clinical practice. EE has been shown to alleviate these effects, prevent damage to hippocampal synaptic plasticity caused by early stress, and improve cognitive function, with positive effects on the central nervous system. EE and physical exercise enhance the plasticity of the adult brain by increasing the expression of neurotrophic factors such as BDNF. These activities can improve cognitive function, reduce anxiety and depressive like behaviors, and promote neuroprotection.
EE has been found to improve the behavior, cellular and molecular defects of animal models of various neurological and NDs, and provide potential new targets for NDs treatment interventions by regulating molecular and cellular mechanisms. For example, EE has been shown to regulate iron metabolism, reduce iron death, and exert neuroprotective effects in models of cerebral ischemia/reperfusion injury [144]. “Mosquito like drug”, an EE mimetic drug, can simulate or enhance the beneficial effects of cognitive activity and physical exercise on the brain. This drug utilizes the therapeutic effects of cognitive stimulation and physical activity to enhance experience dependent plasticity, targeting the molecular and cellular mechanisms of positive effects of EE. This method proposes a new non-invasive treatment strategy to address CNS developmental defects and neurological disorders [145].
Synergy with clinical drugsEE can have a synergistic effect with clinical drugs, improving the treatment effectiveness of NDs. When used in combination with anti-inflammatory drugs, EE can amplify and alleviate neuroinflammatory effects. Dexamethasone, as a key participant in neuroinflammation, is a glucocorticoid with anti-inflammatory effects that can inhibit NF—κ B activation [146]. This type of medication can be used in combination with EE to alleviate drug use and predict phenotypes associated with addiction and stress, significantly reducing inflammation [147]. When combined with neurotrophic factors, EE can promote neurogenesis. BDNF promotes neuronal growth and survival, and EE has been shown to increase BDNF expression. By administering drugs such as 7,8-dihydroxyflavone (7,8-DHF), which are TrkB agonists, this upregulation can be further enhanced, activating the BDNF TrkB signaling cascade and improving cell survival and neuroplasticity [148]. The NF—κ B pathway associated with neuroinflammation can be inhibited by EE, reducing NF—κ B activation. Meanwhile, pathways related to neuroprotection and regeneration, such as the Phosphoinositol-3 kinase (PI3K) Protein Kinase B (Akt) pathway, can be activated by EE to increase BDNF TrkB signaling and promote neuronal growth and survival [148]. This dual regulation leads to a synergistic effect between anti-inflammatory and neuroprotective mechanisms. Targeting the PI3K/Akt and extracellular signal-regulated kinase (ERK) pathways has the potential to inhibit the progression of ND. The disruption of these signaling cascades significantly promotes the pathogenesis of ND, including PD, AD and HD. The Akt pathway regulates neuronal toxicity and survival by interacting with substrates such as FOXos, GSK3β, and caspase-9, typically binding to PI3K. ERK is another key kinase that regulates the proliferation, differentiation, and survival of nerve cells, playing a role in maintaining neuronal health [149].
EE reduces susceptibility to epileptic seizures by enhancing adult neurogenesis in the olfactory cortex (EC) circuit [150]. EE increases hippocampal neurogenesis in adults and significantly reduces susceptibility to epileptic seizures after EE exposure. EE enhanced adult new granulocytes (abDGCs) are activated during epileptic seizures, and the activation of these cells simulates the antiepileptic effect of EE. In addition, whole brain c-Fos mapping showed an increase in the activity of EC CaMKIIα + neurons projected by DG in EE response. These neurons bidirectionally regulate the proliferation and maturation of abDGCs, activate local GABAergic interneurons, and are an important component of EE mediated antiepileptic effects.
In the role of EE in Alzheimer’s disease, EE can promote the conversion of proNGF to mNGF by increasing the level of mature NGF (mNGF), thereby activating the PI3k Akt and MAPK/ERK pathways, promoting neurogenesis, cell survival, cell proliferation, and increased synaptic plasticity [99]. EE also indirectly reduces the activation of the JNK pathway by reducing proNGF levels and directly reducing p75NTR levels, leading to excessive phosphorylation of tau protein, decreased APP phosphorylation, and reduced cell death. EE, along with physical activity, can improve spatial and working memories, and reduce the levels of pathological markers of neurodegeneration such as amyloid beta and tau protein, thereby exerting a synergistic effect on drug therapy. One study investigated the effects of EE and drug intervention (such as metformin) on hippocampal neuronal survival and hippocampal dependent memory in type 2 diabetes rats [151]. The results showed that the combined intervention of EE and EE with metformin improved hippocampal neuron survival and hippocampal dependent memory under stress in T2D rats by enhancing gene expression regulation of neurogenesis and synaptic plasticity. These findings support the view that EE and combined interventions have neuroprotective properties. In a study of AD rats receiving both EE and donepezil (a acetylcholinesterase inhibitor) simultaneously, the performance of the Morris water maze test was significantly improved compared to using EE or donepezil alone or neither [152]. This indicates that EE has the potential to enhance the efficacy of drug therapy.
The drug therapy targets for AD involve multiple key pathophysiological processes, including Aβ aggregation, tau protein hyperphosphorylation, neuroinflammation, oxidative stress, and cholinergic dysfunction. In terms of traditional medicinal plants, such as ginkgo leaves, fir, turmeric, and ginseng, they contain bioactive compounds that can regulate the above targets. For example, flavonoids and terpenoids contained in ginkgo leaves can reduce Aβ deposition and enhance cerebral blood flow, thereby exerting neuroprotective effects. Huperzia serrata is known for its natural source of huperzine A, which can be used as an acetylcholinesterase inhibitor to improve cholinergic function. Turmeric is rich in curcumin, which has anti-inflammatory and antioxidant properties, and can alleviate neuroinflammation and oxidative stress. Ginseng contains ginsenosides, which have shown neuroprotective effects and the ability to counteract amyloidosis [153].
Limitations and challengesAt present, EE still has some limitations in alleviating NDs. Due to the lack of effective biomarkers, the detection efficiency of AD and PD is not high, and currently available biomarkers can often only be diagnosed in the late stages of the disease [154]. This increases the difficulty of disease prevention and limits the effectiveness of early assessment and monitoring of EE. The EE effect is age dependent. In a study, the neuroprotective strategy of EE depended on its early exposure, which limited the effectiveness of EE intervention until later adulthood [155]. In addition, the biomarker detection methods based on cerebrospinal fluid (CSF) and blood also have shortcomings, such as the extremely low concentration of protein biomarkers and the invasive collection process of CSF [154]. These factors affect the application of EE in early disease intervention.
It is difficult to achieve consistency and reproducibility of results through standardization in EE studies and clinical translation. The main obstacle at present is the lack of methods to quantify the degree of EE. Physical motion is considered a key component of EE, but there is currently a lack of evidence to prove a direct relationship between the inanimate and social stimuli generated by EE and the motion induced by EE [156]. All environmental conditions, whether known or unknown, may affect the research results. For example, David et al. found that mice in independent ventilated cages lacking shelter exhibited histological signs of chronic cold stress and altered experimental results (as measured by adrenal gland weight, tumor growth, and adipose tissue) [157].
Even under standardized conditions, differences between laboratories and interactions between genotypes and laboratories may still affect experimental results. The true standardization of all environmental variables is almost impossible to achieve between laboratories, which may result in variations in the results [157]. Moreover, there are challenges in the clinical translation of EE. For outpatient patients, not only is the shared environment diverse, but the non-shared environment is also varied, which is one of the difficulties in EE conversion. In addition, there are technical and feasibility limitations in aligning the feeding conditions in animal experiments with the treatment mode and environment in clinical practice, as well as achieving standardized environmental conditions between hospitals or hospital departments [37].
The use of EE in clinical or experimental settings may result in potential side effects and limitations. Although EE has shown positive effects in the short term, its long-term impact and stability are not fully understood. Some studies suggested that early life experiences led to persistent individualized changes, including behavior, brain plasticity, and epigenetics [158], but the duration of these changes was unknown and might have side effects such as long-term health impacts. Additionally, due to individual differences in response to environmental stimuli, the implementation of EE is limited by individual differences in order to ensure that each subject can achieve optimal results [158].
The emerging treatment strategies currently include the use of stem cell-derived exosomes, which have shown potential to improve cognitive function and reduce A β deposition in preclinical models of Alzheimer’s disease. The PI3K pathway is beneficial for neuronal regeneration after stroke, and activators of this pathway can enhance the survival and differentiation of neuronal cells [159]. Digital therapy (DTx, which may include EE) has limitations in addressing drug non-compliance, adverse reactions, toxicity, and inadequate efficacy of drugs and biologics for chronic diseases. In animal disease models, preclinical studies on EE and other non-pharmacological modes such as physical activity, social interaction, learning, and music have shown improved results and can serve as alternatives to the “active in
Comments (0)