
Remember me
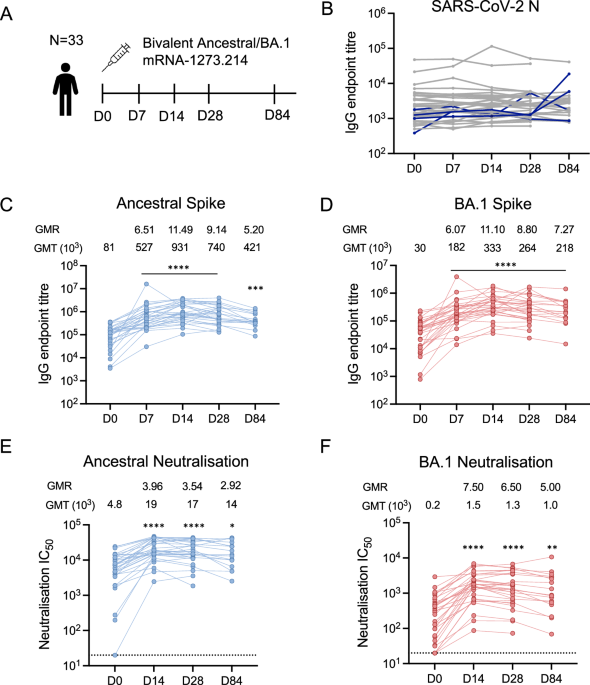
Thirty-three healthy adults (mean age 44.3 years, 16 female/17 male) were administered a booster dose of a bivalent ancestral/BA.1 COVID-19 mRNA vaccine (Fig. 1A, Supplementary Table 1). Antibodies against SARS-CoV-2 nucleocapsid (N) protein were monitored throughout the study to detect subclinical infections not reported by participants. All participants had detectable N-specific antibodies at baseline (Fig. 1B), despite one-third of them (10/33) having no self-reported prior infection. Four participants (depicted as blue lines) had >4-fold rise in N-specific antibodies between consecutive visits, suggesting a breakthrough infection. Consequently, the results of these participants at the impacted time points were excluded from further analysis.
Fig. 1: Antibody responses after bivalent COVID-19 booster vaccination.
A Study design: healthy adults (n = 33) were administered a bivalent mRNA vaccine. Blood samples were collected pre-vaccination day (D)-0, and at days 7, 14, 28 and 84 post-vaccination. B IgG antibody endpoint titres against SARS-CoV-2 Nucleocapsid (N) protein. Individuals with a >4-fold change in titre are indicated in blue. C IgG antibody endpoint titres against Ancestral and D Omicron BA.1 spikes. E Neutralisation titres calculated by 50% virus inhibitory concentration (IC50) against Ancestral and F Omicron BA.1 live viruses. Geometric mean titres (GMT) and geometric mean ratios (GMR) are shown above each graph. For C–F, n = 32 at D0, D7, and D14; n = 31 at D28 and n = 17 at day 84. Asterisk indicates p value of Kruskal–Wallis test with Dunn’s post-test comparing D0 with D7, 14, 28 or 84. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Rapid recall of plasma antibody responses after bivalent booster vaccinationAll participants exhibited baseline binding IgG titres against both ancestral and BA.1 spike. IgG titres increased by day 7, peaked at day 14 and remained significantly elevated compared to baseline at day 84 (p < 0.001; Fig. 1C, D). While geometric mean titres (GMTs) were higher against ancestral than BA.1 spike at all time points, the geometric mean ratios (GMRs) between pre- and post-vaccination time points were similar across the two vaccine antigens (GMR day 14 ancestral = 11.49 vs. BA.1 = 11.10; Fig. 1C, D).
Virus neutralising antibodies (nAbs) are a correlate of protection against symptomatic SARS-CoV-2 infection4. We evaluated nAbs against the two vaccine strains using a live virus microneutralisation assay. The bivalent vaccine increased nAb titres against both viruses at day 14 (Fig. 1E, F), with titres maintained above baseline levels over the 84 days of follow-up (p = 0.012 for ancestral, p = 0.002 for BA.1). While higher nAb GMTs against ancestral virus were observed compared to BA.1 at all time points, GMRs were higher for BA.1 (GMR day 14 BA.1 = 7.19 vs. ancestral = 3.96).
Baseline antigen-specific CD4 T cell frequencies correlate with post-vaccination nAb titresIn contrast to nAb responses, T cells induced by ancestral vaccination or infection are highly cross-reactive with variant spike proteins5,6,7. Nonetheless, a proportion of individuals exhibit comparatively poor recognition of the Omicron spike8. Here, we assessed T-cell responses to the bivalent vaccine using peptide pools covering the mutated regions in the Omicron BA.1 spike and the corresponding regions in the ancestral spike in an ex-vivo T-cell stimulation assay (cell subset definitions and gating shown in Supplementary Fig. 1A). At baseline, frequencies of AIM+ CD4+ T memory (Tmem) cells recognising the BA.1 mutated peptides (median 0.07%) were significantly lower than the frequencies of cells recognising the ancestral (AN) equivalent epitopes (median 0.19%) (2.7-fold difference, p < 0.0001; Fig. 2A). Differences were less pronounced among the AIM+ CD4+ circulating T follicular helper (cTFH) compartment (median 0.07% for AN vs 0.04% for BA.1), while peptide-specific CD8+ Tmem frequencies were low overall (0.03% for AN vs 0.01% for BA.1; Fig. 2A).
Fig. 2: Spike-specific activation-induced marker (AIM) T-cell responses after bivalent COVID-19 booster vaccination.
A Representative staining and frequencies of AIM+ CD4+ T memory (Tmem, non-naïve CXCR5-), CD4+ circulating T follicular helper (cTFH, CD45RA−CXCR5+) cells and CD8+ Tmem cells specific for the mutated epitopes on Omicron BA.1 spike and the corresponding epitopes in the Ancestral spike at baseline. AIM+ cells were defined as OX-40+CD137+ (for both CD4+ subsets) or CD69+CD137+ (for CD8+ T cells). B Representative staining and frequencies of CD8 Tmem recognising ancestral or BA.1 peptide at D0 and D14 post-vaccination. C Frequencies of CD4 Tmem and cTFH recognising ancestral or BA.1 peptides at D0 and D14 post-vaccination. D Correlation between frequencies of BA.1-specific AIM+ CD4+ Tmem at D0 and neutralisation titres against Omicron BA.1 at day 28 post-vaccination. N = 31 for all groups. Bold lines join the median values for each time point. Paired T-cell frequencies were analysed using the Wilcoxon signed-rank test. Correlation was performed by Spearman r test. *p < 0.05, **p < 0.01, ****p < 0.001.
Vaccination drove a significant increase in CD8+ Tmem recognising both ancestral and BA.1 mutated peptides at day 14 (p = 0.013 for AN, p = 0.002 for BA.1; Fig. 2B). Sub-phenotyping based on memory subset revealed that Temra cells (CD45RA+ CCR7−) were the most abundant antigen-specific population, followed by Tem (CD45RA− CCR7−) and then Tcm (CD45RA− CCR7+) subsets, regardless of specificity or timepoint (Supplementary Fig. 1B). Compared to baseline, spike-specific Temra cells were significantly increased at day 14 for both AN and BA.1 (p = 0.013 and 0.001, respectively). Frequencies of antigen-specific CD4+ T cells recognising the ancestral-derived peptides were relatively stable following vaccination (Fig. 2C). In contrast, frequencies of both Tmem and cTFH specific to the BA.1 mutated peptides increased following vaccination (2-fold change for Tmem, p = 0.016, 2.3-fold change for cTFH, p = 0.001; Fig. 2C). Spike-specific CD4 Tmem exhibited a combination of Tcm and Tem phenotypes at both timepoints and regardless of AN or BA.1 specificity (Supplementary Fig. 1C). The increase in BA.1-specific Tmem at day 14 was attributed to a significant increase in the frequency of Tcm (p = 0.004), although there was also a similar trend toward increased frequencies of Tem (p = 0.061). Collectively, bivalent vaccination boosted both CD4 and CD8 T cell responses toward novel epitopes present in the BA.1 spike protein.
Previous studies have suggested that pre-vaccination CD4 T cell memory can predict post-vaccination antibody responses9. We, therefore, assessed whether spike-specific T-cell responses can predict nAb titres following bivalent vaccination. Interestingly, frequencies of BA.1-specific CD4+ Tmem at baseline correlated with BA.1 nAb titres at 28 days post-vaccination (Spearman r = 0.57, p = 0.0008; Fig. 2D), suggesting a role for T cell help and/or prior exposure to BA.1 in determining post-booster neutralising titres.
Bivalent booster vaccination predominantly drives activation and expansion of memory B cells cross-recognising ancestral and BA.1 spikeMemory B cells provide a mechanism for rapid supplementation of protective immunity upon re-infection or vaccination via differentiation into antibody-secreting cells. We assessed spike-specific B cell (CD19+ IgD−) responses using flow cytometry and probes of fluorochrome-conjugated ancestral and Omicron BA.1 spike proteins (gating shown in Supplementary Fig. 2). Three B-cell populations could be resolved: those binding the ancestral spike (AN+ only), binding Omicron BA.1 spike (BA.1+ only) or those cross-reactive to both (AN+ BA.1+; Fig. 3A). Following bivalent vaccination, a significant increase in the frequencies of AN+ BA.1+ cross-reactive B cells was observed (p < 0.001), in contrast to the minimal change in frequency of AN+ only or BA.1+ mono-specific populations (Fig. 3B). Post-vaccination frequencies of BA.1+ cells were generally low in all individuals (median 0.023%), while most subjects had clear populations of AN+ (median 0.31%) or AN+ BA.1+ (median 0.58%) B cells.
Fig. 3: Spike-specific B-cell responses after bivalent COVID-19 booster vaccination.
A Representative staining of antigen-specific memory B cells (CD19+IgD−) at baseline and 28 days post-vaccination. Two fluorochrome-conjugated spike probes were used to differentiate B-cell populations that were either specific for Ancestral (AN only), Omicron BA.1 (BA.1 only) or both spike proteins (AN+BA.1+). B Frequencies of antigen (Ag)-specific IgD− CD19+ B cells at days 0 and 28. C Proportion of IgG+ (solid bars), IgA+ (open bars), and IgM+ (checked bars) cells in AN, BA.1, or AN+BA.1+ MBC populations. D Proportion of activated CD27+ CD21− cells among MBC populations. E Correlation between baseline frequencies of AN+BA.1+ B cells and neutralisation titres against ancestral and Omicron BA.1 viruses at D28. N = 31 for all groups. Bold lines join the medians for each group. B-cell frequencies were analysed by the Wilcoxon signed-rank test. Correlations were analysed by Spearman r test. ***p < 0.001, ****p < 0.0001.
We next investigated the distribution of immunoglobulin isotypes and the activation status of spike-specific B cells over the course of vaccination. In both pre- and post-vaccination, all spike antigen-specific MBC populations were predominantly IgG, with no substantial changes between day 0 and 28 (Fig. 3C). To identify activated B cells, we assessed the frequency of CD27+ memory B cells that downregulated expression of CD21. In response to bivalent booster immunisation, we observed increased proportions of CD27+CD21- cells in all three spike-specific B cell populations, with the most consistent activation in AN+ BA.1+ cells (median 3.16% at day 0 vs 15.30% at day 28, p < 0.0001; Fig. 3D).
Finally, we sought to determine whether the magnitude of the baseline MBC pool was associated with the post-vaccine serological response. In contrast to the relationship between baseline CD4 T cell memory and nAb titres, there was only a weak association between pre-vaccination cross-reactive B cell frequencies (p = 0.054 for ancestral virus, p = 0.06 for BA.1; Fig. 3E). No correlation was found between nAb titre and the frequency of either AN+ only or BA.1+ only populations.
Vaccine-induced antibodies and B cells cross-react to Omicron XBB subvariantShortly after the bivalent vaccine roll-out in Australia (September 2022), the previously circulating BA.1 variant was replaced with XBB (lineages XBB.1.16, XBB.1.5, and XBB.1.9), which circulated widely until late 2023. Hence, we assessed antibodies and B cells generated by the bivalent ancestral and Omicron BA.1 booster vaccine for cross-reactivity against XBB.1.5. We found an increase in IgG antibodies binding to the XBB.1.5 spike as early as 7 days after vaccination, with GMTs and GMRs peaking at 14 days post-vaccination (Fig. 4A). At baseline, 15/33 participants had detectible neutralising activity against live XBB virus (Fig. 4B). Vaccination elicited nAb in 91% of participants, with only 3 individuals failing to develop detectable XBB-specific nAb at either 14- or 28-days post-vaccination (Fig. 4B). XBB nAb titres remained significantly elevated at day 84 of follow-up compared to baseline (p = 0.0032; Fig. 4B), although at all timepoints titres against XBB remained markedly lower than against ancestral and BA.1 viruses (Figs. 4B and 1E, F).
Fig. 4: Cross-reactive antibody and B-cell responses to Omicron XBB after bivalent booster vaccination.
A IgG endpoint titres and (B) live virus IC50 titres against XBB spike. The dotted line indicates the limit of detection. Geometric mean titres (GMT) and geometric mean ratios (GMR) are shown above the graph. Asterisk indicates p value of Kruskal–Wallis test with Dunn’s post-test comparing D0 with D7, 14, 28 or 84. C Representative staining of XBB.1.5 spike probe among populations of AN+, AN+BA.1+ or AN-BA.1− IgG+ memory B cells. D Proportion of XBB+ cells in Ag+ IgG+ B cells at baseline and day 28 post-vaccination. Bold lines join the median for each time point. N = 32 at days 0 and 14; N = 31 at day 28 and N = 17 at day 84. B-cell proportions were analysed using a non-parametric paired-sample Wilcoxon signed-rank test. E Correlation between frequencies of BA.1-specific AIM+ CD4+ Tmem or F frequencies of AN+BA.1+XBB.1.5+ cross-reactive MBC at D0 and neutralisation titres against XBB at D28 post-vaccination (n = 31). Correlation assessed by Spearman r test. **p < 0.01, ***p < 0.001, ****p < 0.0001.
To identify MBC populations with cross-reactivity to the XBB.1.5 spike, we examined the extent of XBB.1.5 probe binding within AN+ only, BA.1+ only, AN+ BA.1+ or AN-BA.1- IgG+ B-cell populations (Fig. 4C). The vast majority of XBB.1.5 recognition occurred within the AN+ BA.1+ MBC compartment, with minimal cross-reactivity between XBB.1.5 and AN+ only cells, and sporadic XBB.1.5 binding within the minor BA.1+ only subset (Fig. 4D). Vaccination had a modest, but significant, impact on the proportion of XBB-binding cells within the AN+BA.1+ population (Fig. 4D; median 62.1% at day 0 versus 69.8% at day 28; p < 0.001).
Given the relationship between vaccine-strain nAb titres and CD4 T cell memory (Fig. 2D), we performed a similar correlation for XBB nAb titres at D28 with either baseline BA.1-specific CD4 Tmem (Fig. 4E) or baseline AN+BA.1+XBB+ MBC (Fig. 4F). Once again, baseline CD4 T cell memory correlated with post-vaccine nAb (p = 0.003, r = 0.52), while no such relationship was present for cross-reactive MBC (p = 0.13, r = 0.28).
Predictors of vaccine-elicited neutralisation against antigenic variant virusesConsidering the broad cross-reactivity of spike-specific memory B cells, it is unsurprising that recipients of the bivalent mRNA vaccine showed expansion of serological responses able to recognise variant XBB spike. Nevertheless, the wide variation in serum XBB neutralising activity between participants suggests that booster immunisation may have provided differential protection against XBB viruses within vaccine recipients. To better understand the demographic and immunologic factors that influence post-vaccination neutralising titres, we constructed a multiple regression model. The model took into account participant age, gender, number of prior COVID-19 vaccinations, number of self-reported prior infections, days since last antigen exposure (either vaccination or infection), baseline IgG titres against Ancestral or BA.1 spike, baseline nAb titres against ancestral, BA.1 or XBB virus, baseline frequencies of Ancestral, BA.1 or cross-reactive memory B cells, and baseline frequencies of AIM+ CD4+ cTFH or Tmem.
Post-vaccination nAb titres against either ancestral or BA.1 virus were best predicted by a combination of baseline ancestral nAb titres and baseline ancestral spike-specific CD4 Tmem frequencies (Table 1). We next constructed a similar model to assess the variables that predict the post-vaccination nAb titre against XBB. Once again, baseline ancestral nAb titres and baseline spike-specific CD4 Tmem frequencies were found to be the only significant independent predictors of XBB nAbs (Table 1), suggesting that, in this cohort, a combination of pre-existing CD4 T cell help and prior antibody responses to the ancestral strain were the strongest predictors of post-vaccination antibody breadth against a future viral variant.
Table 1 Significant predictors of post-vaccination titres
Comments (0)